Glavne stopnje evolucije možganov pri vretenčarjih. Glavne stopnje evolucije možganov pri vretenčarjih po M.A.
V evoluciji je živčni sistem prestal več stopenj razvoja, ki so postale prelomnice v kvalitativni organizaciji njegovih dejavnosti. Te stopnje se razlikujejo po številu in vrstah nevronskih tvorb, sinaps, znakih njihove funkcionalne specializacije in oblikovanju skupin nevronov, ki so med seboj povezane s skupnimi funkcijami. Obstajajo tri glavne stopnje strukturne organizacije živčnega sistema: difuzna, nodularna, cevasta.
DifuznoŽivčni sistem je najstarejši, najdemo ga pri coelenteratah (hidra). Za takšen živčni sistem je značilna množica povezav med sosednjimi elementi, kar omogoča, da se vzbujanje prosto širi po živčnem omrežju v vse smeri.
Ta vrsta živčnega sistema zagotavlja široko zamenljivost in s tem večjo zanesljivost delovanja, vendar so te reakcije nenatančne in nejasne.
Vozlišče vrsta živčnega sistema je značilna za črve, mehkužce in rake.

Zanj je značilno, da so povezave živčnih celic organizirane na določen način, vzbujanje poteka po strogo določenih poteh. Ta organizacija živčnega sistema se izkaže za bolj ranljivo. Poškodba enega vozla povzroči motnje v delovanju celotnega organizma kot celote, vendar so njegove lastnosti hitrejše in natančnejše.
cevastoŽivčni sistem je značilen za hordate; vključuje značilnosti difuznega in nodularnega tipa. Živčni sistem višjih živali je vzel vse najboljše: visoko zanesljivost difuznega tipa, natančnost, lokalnost, hitrost organizacije reakcij nodalnega tipa.
Vodilna vloga živčnega sistema
Na prvi stopnji razvoja sveta živih bitij je interakcija med najpreprostejšimi organizmi potekala skozi vodno okolje primitivnega oceana, v katerega so vstopile kemične snovi, ki so jih sproščali. Prva najstarejša oblika interakcije med celicami večceličnega organizma je kemična interakcija preko presnovnih produktov, ki vstopajo v telesne tekočine. Takšni presnovni produkti ali metaboliti so produkti razgradnje beljakovin, ogljikovega dioksida itd. To je humoralni prenos vplivov, humoralni mehanizem korelacije ali povezave med organi.
Za humoralno povezavo so značilne naslednje značilnosti:
- pomanjkanje točnega naslova, kamor se pošlje kemična snov, ki vstopi v kri ali druge telesne tekočine;
- kemikalija se širi počasi;
- kemikalija deluje v majhnih količinah in se običajno hitro razgradi ali izloči iz telesa.
Humorne povezave so skupne tako živalskemu kot rastlinskemu svetu. Na določeni stopnji razvoja živalskega sveta se v povezavi s pojavom živčevja oblikuje nova, živčna oblika povezav in regulacije, ki kakovostno loči živalski svet od rastlinskega. Višja kot je razvitost živalskega organizma, večja je vloga medsebojnega delovanja organov skozi živčni sistem, ki ga imenujemo refleks. V višjih živih organizmih živčni sistem uravnava humoralne povezave. Za razliko od humoralne ima živčna povezava točno določeno smer do določenega organa in celo skupine celic; komunikacija poteka stokrat hitreje od hitrosti distribucije kemikalij. Prehoda iz humoralne povezave v živčno povezavo ni spremljalo uničenje humoralne povezave med celicami telesa, temveč podreditev živčnih povezav in nastanek nevrohumoralnih povezav.
Na naslednji stopnji razvoja živih bitij se pojavijo posebni organi - žleze, v katerih nastajajo hormoni, ki nastanejo iz hranilnih snovi, ki vstopajo v telo. Glavna funkcija živčnega sistema je uravnavanje delovanja posameznih organov med seboj in v interakciji telesa kot celote z zunanjim okoljem. Vsak vpliv zunanjega okolja na telo se pojavi predvsem na receptorjih (čutilnih organih) in se izvaja s spremembami, ki jih povzročajo zunanje okolje in živčni sistem. Ko se živčni sistem razvija, njegov najvišji del – možganske hemisfere – postane »upravitelj in razdelilec vseh telesnih dejavnosti«.
Zgradba živčnega sistema
Živčni sistem tvori živčno tkivo, ki je sestavljeno iz ogromne količine nevroni- živčna celica s procesi.

Živčni sistem je običajno razdeljen na centralni in periferni.

Centralni živčni sistem vključuje možgane in hrbtenjačo ter periferni živčni sistem- živci, ki segajo iz njih.

Možgani in hrbtenjača so skupek nevronov. V prerezu možganov ločimo belo in sivo snov. Sivo snov sestavljajo živčne celice, belo snov pa sestavljajo živčna vlakna, ki so odrastki živčnih celic. V različnih delih centralnega živčnega sistema je lokacija bele in sive snovi drugačna. V hrbtenjači se siva snov nahaja znotraj, bela snov pa zunaj; v možganih (možganske hemisfere, mali možgani), nasprotno, siva snov je zunaj, bela snov pa znotraj. V različnih delih možganov obstajajo ločeni skupki živčnih celic (siva snov), ki se nahajajo znotraj bele snovi - jedrca. Skupki živčnih celic se nahajajo tudi zunaj centralnega živčnega sistema. Imenujejo se vozlišča in spadajo v periferni živčni sistem.
Refleksna aktivnost živčnega sistema
Glavna oblika delovanja živčnega sistema je refleks. Refleks- reakcija telesa na spremembe v notranjem ali zunanjem okolju, ki se izvaja s sodelovanjem centralnega živčnega sistema kot odgovor na draženje receptorjev.
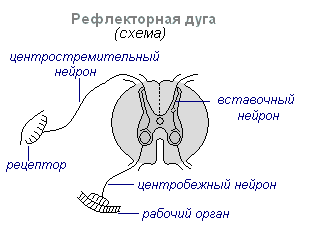
Pri vsakem draženju se vzbujanje iz receptorjev prenaša po centripetalnih živčnih vlaknih v centralni živčni sistem, od koder skozi interneuron vzdolž centrifugalnih vlaken gre na obrobje do enega ali drugega organa, katerega aktivnost se spremeni. Ta celotna pot skozi centralni živčni sistem do delovnega organa se imenuje refleksni lok običajno tvorijo trije nevroni: senzorični, interkalarni in motorični. Refleks je kompleksno dejanje, pri katerem sodeluje bistveno večje število nevronov. Vzbujanje, ki vstopi v centralni živčni sistem, se razširi na številne dele hrbtenjače in doseže možgane. Zaradi interakcije številnih nevronov se telo odzove na draženje.
Hrbtenjača
Hrbtenjača- vrvica dolžine približno 45 cm, premera 1 cm, ki se nahaja v hrbteničnem kanalu, prekrita s tremi možganskimi ovojnicami: dura, arahnoidna in mehka (vaskularna).
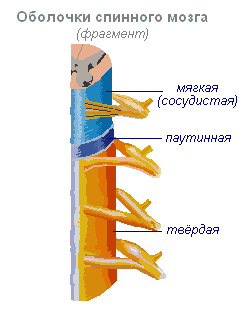
Hrbtenjača se nahaja v hrbteničnem kanalu in je vrvica, ki na vrhu prehaja v podolgovato medullo, spodaj pa se konča na ravni drugega ledvenega vretenca. Hrbtenjača je sestavljena iz sive snovi, ki vsebuje živčne celice, in bele snovi, ki je sestavljena iz živčnih vlaken. Siva snov se nahaja znotraj hrbtenjače in je z vseh strani obdana z belo snovjo.
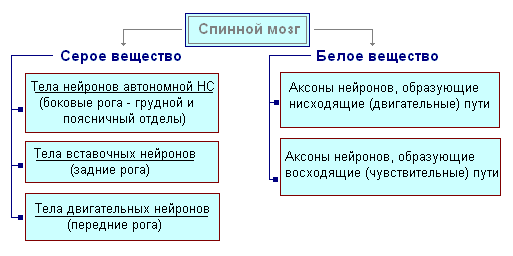
V prerezu je siva snov podobna črki H. Razlikuje sprednji in zadnji rog ter povezovalno prečko, v središču katere je ozek kanal hrbtenjače, ki vsebuje cerebrospinalno tekočino. V prsnem predelu so stranski rogovi. Vsebujejo telesa nevronov, ki inervirajo notranje organe. Belo snov hrbtenjače tvorijo živčni procesi. Kratki procesi povezujejo dele hrbtenjače, dolgi pa tvorijo prevodni aparat dvostranskih povezav z možgani.
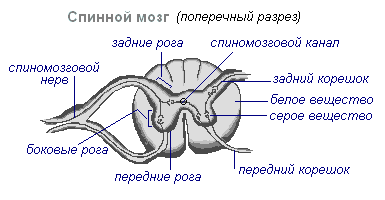
Hrbtenjača ima dve zgostitvi - vratno in ledveno, iz katerih segajo živci do zgornjih in spodnjih okončin. Iz hrbtenjače izhaja 31 parov hrbtenjačnih živcev. Vsak živec se začne iz hrbtenjače z dvema koreninama - sprednjo in zadnjo. Zadnje korenine - občutljiva sestavljajo procesi centripetalnih nevronov. Njihova telesa se nahajajo v hrbteničnih ganglijih. Sprednje korenine - motor- so procesi centrifugalnih nevronov, ki se nahajajo v sivi snovi hrbtenjače. Zaradi zlitja sprednjih in zadnjih korenin nastane mešani hrbtenični živec. Hrbtenjača vsebuje centre, ki uravnavajo najpreprostejše refleksne akte. Glavne funkcije hrbtenjače so refleksna aktivnost in prevajanje vzbujanja.

Človeška hrbtenjača vsebuje refleksne centre za mišice zgornjih in spodnjih okončin, znojenje in uriniranje. Funkcija vzbujanja je, da impulzi iz možganov v vsa področja telesa in nazaj prehajajo skozi hrbtenjačo. Centrifugalni impulzi iz organov (kože, mišic) se po ascendentnih poteh prenašajo v možgane. Po padajočih poteh se centrifugalni impulzi prenašajo iz možganov v hrbtenjačo, nato na periferijo, v organe. Ko so poti poškodovane, pride do izgube občutljivosti v različnih delih telesa, kršitve prostovoljnih mišičnih kontrakcij in sposobnosti gibanja.
Evolucija možganov vretenčarjev
Tvorba osrednjega živčevja v obliki nevralne cevi se najprej pojavi pri hordatih. U nižji hordati nevralna cev traja vse življenje, višji- vretenčarji - v embrionalnem stadiju se na hrbtni strani oblikuje nevralna plošča, ki se zarije pod kožo in se zvije v cev. V embrionalni fazi razvoja nevralna cev tvori v sprednjem delu tri otekline - tri možganske vezikle, iz katerih se razvijejo deli možganov: sprednji vezikel daje prednji možgan in diencefalon, srednji vezikel se spremeni v srednje možgane, zadnji vezikel tvori male možgane in podolgovato medullo. Teh pet predelov možganov je značilnih za vse vretenčarje.
Za nižji vretenčarji- ribe in dvoživke - za katere je značilna prevlada srednjih možganov nad drugimi deli. U dvoživke Sprednji možgani se nekoliko povečajo in na strehi hemisfer nastane tanka plast živčnih celic - primarni medularni obok, starodavna skorja. U plazilci Prednji del možganov se znatno poveča zaradi kopičenja živčnih celic. Večino strehe hemisfer zavzema starodavna skorja. Pri plazilcih se prvič pojavi zametek nove skorje. Hemisfere prednjih možganov se plazijo na druge dele, zaradi česar nastane ovinek v predelu diencefalona. Od starodavnih plazilcev so možganske hemisfere postale največji del možganov.
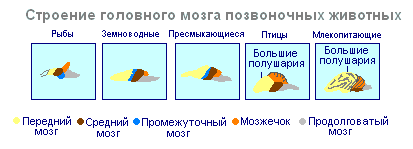
V strukturi možganov ptice in plazilci veliko skupnega. Na strehi možganov je primarna skorja, srednji možgani so dobro razviti. Vendar se pri pticah v primerjavi s plazilci skupna masa možganov in relativna velikost prednjih možganov povečata. Mali možgani so veliki in imajo nagubano strukturo. U sesalci prednji možgani dosežejo največjo velikost in kompleksnost. Večino možganske snovi sestavlja neokorteks, ki služi kot središče višjega živčnega delovanja. Vmesni in srednji del možganov pri sesalcih sta majhna. Širječe hemisfere prednjih možganov jih pokrivajo in stiskajo pod seboj. Nekateri sesalci imajo gladke možgane brez utorov ali vijug, večina sesalcev pa ima utore in vijuge v možganski skorji. Pojav žlebov in konvolucij se pojavi zaradi rasti možganov z omejenimi dimenzijami lobanje. Nadaljnja rast skorje vodi do pojava zlaganja v obliki utorov in zvitkov.
možgani
Če je hrbtenjača pri vseh vretenčarjih bolj ali manj enako razvita, potem se možgani pri različnih živalih bistveno razlikujejo po velikosti in kompleksnosti strukture. Sprednji možgani so med evolucijo podvrženi posebno dramatičnim spremembam. Pri nižjih vretenčarjih so prednji možgani slabo razviti. Pri ribah ga predstavljajo vohalni režnji in jedra sive snovi v debelini možganov. Intenziven razvoj prednjih možganov je povezan s prihodom živali na kopno. Loči se na diencefalon in dve simetrični polobli, ki se imenujeta telencefalon. Siva snov na površini prednjih možganov (skorja) se najprej pojavi pri plazilcih, razvija se naprej pri pticah in predvsem pri sesalcih. Resnično velike poloble sprednjih možganov postanejo le pri pticah in sesalcih. Pri slednjem pokrivajo skoraj vse druge dele možganov.
Možgani se nahajajo v lobanjski votlini. Vključuje možgansko deblo in telencefalon (možgansko skorjo).

Možgansko deblo sestavljajo medula oblongata, pons, srednji možgani in diencefalon.
Medulla oblongata je neposredno nadaljevanje hrbtenjače in, ko se širi, prehaja v zadnje možgane. V bistvu ohranja obliko in strukturo hrbtenjače. V debelini podolgovate medule so kopičenja sive snovi - jedra kranialnih živcev. Zadnja os vključuje mali možgani in pons. Mali možgani se nahajajo nad podolgovato medullo in imajo kompleksno zgradbo. Na površini hemisfer malih možganov siva snov tvori skorjo, znotraj malih možganov pa njena jedra. Tako kot podolgovata medula opravlja dve funkciji: refleksno in prevodno. Vendar pa so refleksi podolgovate medule bolj zapleteni. To se odraža v njegovem pomenu pri uravnavanju srčne dejavnosti, stanja krvnih žil, dihanja in potenja. Centri vseh teh funkcij se nahajajo v medulli oblongati. Tu so centri za žvečenje, sesanje, požiranje, slino in želodčni sok. Kljub svoji majhnosti (2,5–3 cm) je medula oblongata vitalni del centralnega živčnega sistema. Poškodba le-tega lahko povzroči smrt zaradi prenehanja dihanja in delovanja srca. Prevodniška funkcija medule oblongate in mosta je prenos impulzov iz hrbtenjače v možgane in nazaj.
IN srednji možgani nahajajo se primarni (subkortikalni) centri vida in sluha, ki izvajajo refleksne orientacijske reakcije na svetlobno in zvočno stimulacijo. Te reakcije se izražajo v različnih gibih trupa, glave in oči proti dražljajem. Srednji možgani so sestavljeni iz cerebralnih pecljev in kvadrigeminalisa. Srednji možgani uravnavajo in razporejajo tonus (napetost) skeletnih mišic.
diencefalon sestavljata dva oddelka - talamus in hipotalamus, od katerih je vsaka sestavljena iz velikega števila jeder vidnega talamusa in subtalamične regije. Skozi vidne griče se centripetalni impulzi prenašajo v možgansko skorjo iz vseh receptorjev v telesu. Noben centripetalni impulz, ne glede na to, od kod prihaja, ne more preiti v skorjo, mimo vizualnih gričev. Tako prek diencefalona vsi receptorji komunicirajo z možgansko skorjo. V subtuberkularnem predelu so centri, ki vplivajo na metabolizem, termoregulacijo in endokrine žleze.
Mali možgani ki se nahaja za podolgovato medullo. Sestavljen je iz sive in bele snovi. Vendar se za razliko od hrbtenjače in možganskega debla siva snov – skorja – nahaja na površini malih možganov, bela snov pa se nahaja znotraj, pod skorjo. Mali možgani usklajujejo gibe, jih naredijo jasne in gladke, igrajo pomembno vlogo pri ohranjanju ravnotežja telesa v prostoru in vplivajo tudi na mišični tonus. Ko so mali možgani poškodovani, oseba doživi zmanjšanje mišičnega tonusa, motnje gibanja in spremembe v hoji, govor se upočasni itd. Vendar pa se čez nekaj časa obnovi gibanje in mišični tonus zaradi dejstva, da nedotaknjeni deli centralnega živčnega sistema prevzamejo funkcije malih možganov.
Velike hemisfere- največji in najbolj razvit del možganov. Pri človeku tvorijo glavnino možganov in so prekrite s skorjo po celotni površini. Siva snov prekriva zunanjost polobel in tvori možgansko skorjo. Človeška možganska skorja je debela od 2 do 4 mm in je sestavljena iz 6–8 plasti, ki jih tvori 14–16 milijard celic, različnih po obliki, velikosti in funkcijah. Pod skorjo je bela snov. Sestavljen je iz živčnih vlaken, ki povezujejo skorjo z nižjimi deli centralnega živčnega sistema in posamezne režnje hemisfer med seboj.
Možganska skorja ima vijuge, ločene z utori, ki znatno povečajo njeno površino. Trije najgloblji utori delijo hemisfere na režnje. Vsaka polobla ima štiri režnje: frontalni, parietalni, temporalni, okcipitalni. Vzbujanje različnih receptorjev vstopi v ustrezna zaznavna področja korteksa, imenovana cone, od tu pa se prenesejo na določen organ in ga spodbudijo k delovanju. V skorji se razlikujejo naslednje cone. Slušno območje ki se nahaja v temporalnem režnju, sprejema impulze iz slušnih receptorjev.

Vizualno območje leži v okcipitalnem predelu. Sem prihajajo impulzi iz očesnih receptorjev.

Vohalna cona nahaja se na notranji površini temporalnega režnja in je povezan z receptorji v nosni votlini.

Senzorično-motorični območje se nahaja v čelnem in parietalnem režnju. To območje vsebuje glavne centre gibanja nog, trupa, rok, vratu, jezika in ustnic. Tu je tudi središče govora.

Možganske hemisfere so najvišji del osrednjega živčnega sistema, ki nadzoruje delovanje vseh organov pri sesalcih. Pomen možganskih hemisfer pri človeku je tudi v tem, da predstavljajo materialno osnovo duševne dejavnosti. I. P. Pavlov je pokazal, da duševna dejavnost temelji na fizioloških procesih, ki se odvijajo v možganski skorji. Mišljenje je povezano z delovanjem celotne možganske skorje in ne le z delovanjem posameznih njenih področij.
| Oddelek za možgane | Funkcije | |
| Medulla oblongata | Dirigent | Povezava med hrbtenico in ležečimi deli možganov. |
| Refleks | Regulacija dihalnega, kardiovaskularnega, prebavnega sistema:
|
|
| Pons | Dirigent | Povezuje hemisfere malih možganov med seboj in z možgansko skorjo. |
| Mali možgani | Usklajevanje | Koordinacija prostovoljnih gibov in ohranjanje položaja telesa v prostoru. Regulacija mišičnega tonusa in ravnovesja |
| Srednji možgani | Dirigent | Približni refleksi na vizualne in zvočne dražljaje ( obrača glavo in telo). |
| Refleks |
|
|
| diencefalon | talamus
hipotalamus
|
|
Možganska skorja
Površina možganska skorja pri človeku je približno 1500 cm 2, kar je večkrat več od notranje površine lobanje. Ta velika površina skorje je nastala zaradi razvoja velikega števila utorov in zvitkov, zaradi česar je večina skorje (približno 70%) koncentrirana v utorih. Največji utori možganskih hemisfer so osrednji, ki poteka čez obe polobli, ter časovno, ki ločuje temporalni reženj od ostalih. Možganska skorja ima kljub majhni debelini (1,5–3 mm) zelo zapleteno strukturo. Ima šest glavnih plasti, ki se razlikujejo po zgradbi, obliki in velikosti nevronov ter povezav. V korteksu so središča vseh čutnih (receptorskih) sistemov, predstavniki vseh organov in delov telesa. V zvezi s tem se centripetalni živčni impulzi iz vseh notranjih organov ali delov telesa približajo skorji in lahko nadzorujejo njihovo delo. Skozi možgansko skorjo se zaprejo pogojni refleksi, s pomočjo katerih se telo nenehno, vse življenje, zelo natančno prilagaja spreminjajočim se pogojem obstoja, okolju.
Državna izobraževalna ustanova višjega strokovnega izobraževanja "Stavropol State Medical Academy" Ministrstva za zdravje in socialni razvoj Ruske federacijeOddelek za biologijo z ekologijo
A.B. Khojayan
N.N. Fedorenko
L.A. Krasnova
O NEKATERIH VPRAŠANJIH EVOLUCIJE
(dodano)
Metodološki priročnik za študente 1. letnika StSMA
STAVROPOL,
UDK 57:575.
O nekaterih vprašanjih evolucije. Metodološki priročnik za študente 1. letnika. Založnik: StSMA. 2009 str.31.
V učbeniku biologije, ur. A.A.Slyusareva in S.V. Žukova, ki ga uporabljajo študenti 1. letnika pri študiju medicinske biologije in genetike, nekatera vprašanja teorije evolucije zahtevajo dopolnitev in pojasnilo. Osebje Oddelka za biologijo St. Državne medicinske akademije je menilo, da je treba sestaviti ta metodološki priročnik o nekaterih vprašanjih teorije evolucije žive narave.
Sestavil: Doktor medicinskih znanosti, prof. Khojayan A.B.,
Kandidat medicinskih znanosti, izredni profesor Fedorenko N.N.,
Kandidat medicinskih znanosti, izredni profesor Krasnova L.A.
© Država Stavropol
Medicinska akademija, 2009
^ FILOGENEZA ORGANSKIH SISTEMOV PRI ŽIVALIH
Osnov zgradbe in delovanja različnih organov in organskih sistemov pri živalih in ljudeh ni mogoče dovolj in v celoti razumeti brez poznavanja njihovega zgodovinskega nastanka, to je filogeneze.
^ Filogenija živčnega sistema .
Vsi živi organizmi skozi svoje življenje doživljajo različne vplive iz zunanjega okolja, na katere se odzivajo s spremembo vedenja ali fizioloških funkcij. Ta sposobnost odzivanja na vplive okolja se imenuje razdražljivost.
Razdražljivost se pojavlja že pri praživalih in se izraža v spremembah njihovih vitalnih procesov ali vedenja kot odziv na dražljaje, kot so kemikalije, temperatura in svetloba.
Pri večceličnih živalih se pojavi poseben sistem celic - nevroni, ki se lahko na določene dražljaje odzovejo z živčnim impulzom, ki ga prenašajo na druge celice telesa. Zbirka živčnih celic tvori živčni sistem, katerega kompleksnost zgradbe in delovanja narašča s kompleksnostjo organizacije živali. Glede na slednje so se pri večceličnih živalih v evoluciji razvili trije glavni tipi živčnega sistema: retikularni (difuzni), ganglijski (nodularni) in cevasti.
^
D  razpršeno (mrežno)
)
živčen
sistem je značilen za najbolj primitivne živali - coelenterate. Njihov živčni sistem sestavljajo nevroni, razpršeni po telesu, ki se s svojimi procesi stikajo med seboj in s celicami, ki jih inervirajo, ter tvorijo nekakšno mrežo. Takšna organizacija živčnega sistema zagotavlja visoko zamenljivost nevronov in s tem večjo zanesljivost delovanja. Vendar so odzivi s to vrsto organizacije živčnega sistema nenatančni in nejasni.
razpršeno (mrežno)
)
živčen
sistem je značilen za najbolj primitivne živali - coelenterate. Njihov živčni sistem sestavljajo nevroni, razpršeni po telesu, ki se s svojimi procesi stikajo med seboj in s celicami, ki jih inervirajo, ter tvorijo nekakšno mrežo. Takšna organizacija živčnega sistema zagotavlja visoko zamenljivost nevronov in s tem večjo zanesljivost delovanja. Vendar so odzivi s to vrsto organizacije živčnega sistema nenatančni in nejasni.
^ Nodularni (ganglijski) tip je naslednji korak v razvoju živčnega sistema. Značilen je za vse črve, iglokožce, mehkužce in členonožce. Imajo koncentracijo nevronskih teles v obliki posameznih skupkov – vozlov (ganglijev). Poleg tega so pri ploščatih in okroglih črvih takšni vozli le na sprednjem koncu telesa, kjer so koncentrirani organi za zajemanje hrane in čutila. Pri obročkih in členonožcih, katerih telo je razdeljeno na segmente, razen ganglijev glave, se oblikuje trebušna veriga živčnih ganglijev, ki uravnavajo delovanje tkiv in organov določenega segmenta (annelis) ali skupine segmentov (členonožci). ). Vedno pa ostaja najbolj razvit glavni ganglij, ki je koordinacijski in regulacijski center v odnosu do ostalih ganglijev. Za to vrsto živčnega sistema je značilna določena organizacija: kjer vzbujanje poteka strogo po določeni poti, kar daje prednost pri hitrosti in natančnosti reakcije. Toda ta vrsta živčnega sistema je zelo ranljiva.
Za hordate je značilno cevasto vrsta živčnega sistema. V embrionalnem obdobju se iz ektoderma nad notohordom oblikuje nevralna cev, ki v suličniku ostane vse življenje in služi kot osrednji del živčnega sistema, pri vretenčarjih pa se spremeni v hrbtenjačo in možgane. V tem primeru se možgani razvijejo iz sprednjega dela nevralne cevi in iz preostalega dela hrbtenjače.
Možgani pri vretenčarjih so sestavljeni iz petih delov: prednjih možganov, srednjih srednjih možganov, podolgovate medule in malih možganov.

^ EVOLUCIJA MOŽGANOV PRI VRETENČARJIH
Nastajanje možganov pri zarodkih vseh vretenčarjev se začne s pojavom oteklin - možganskih veziklov - na sprednjem koncu nevralne cevi. Najprej so trije, nato pa pet. Iz prednjih možganov se nato oblikujejo prednji možgani in diencefalon, iz srednjega - mezencefalon, iz zadnjega pa - mali možgani in podolgovata medula. Slednji brez ostre meje prehaja v hrbtenjačo
V nevralni cevi je votlina - neurocoel, ki med nastankom petih možganskih veziklov tvori podaljške - možganske prekate (v teh predelih možganov so 4) dno (baza) in a ločimo streho (plašč). Streha se nahaja zgoraj - in dno se nahaja pod ventrikli.
Možganska snov je heterogena – predstavljata jo siva in bela snov. Siva je skupek nevronov, bela pa nastane iz odrastkov nevronov, prekritih z maščobo podobno snovjo (mielinska ovojnica), ki daje možganski snovi belo barvo. Plast sive snovi na strehi katerega koli dela možganov se imenuje skorja.
Čutilni organi igrajo pomembno vlogo pri razvoju živčnega sistema. Koncentracija čutnih organov na sprednjem koncu telesa je bila tista, ki je določila progresivni razvoj glave nevralne cevi. Menijo, da je sprednji možganski mehurček nastal pod vplivom vohalnih, srednjih - vizualnih in posteriornih - slušnih receptorjev.
^ Ribi
Prednji možgani majhna, ni razdeljena na poloble, ima samo en ventrikel. Njena streha ne vsebuje živčnih elementov, ampak jo tvori epitelij. Nevroni so koncentrirani na dnu ventrikla v striatumu in v vohalnih režnjih, ki se raztezajo pred sprednjimi možgani. V bistvu prednji možgani delujejo kot vohalni center.
^ Srednji možgani je najvišje regulativno in integrativno središče. Sestavljen je iz dveh optičnih režnjev in je največji del možganov. Ta vrsta možganov, kjer je najvišji regulacijski center srednji možgan, se imenuje ihtiopsidpim.
^ diencefalon sestavljena iz strehe (talamus) in dna (hipotalamus) je povezana s hipotalamusom, pinealna žleza pa je povezana s talamusom.
Mali možgani pri ribah je dobro razvit, saj so njihova gibanja zelo raznolika.
^ Medulla oblongata brez ostre meje prehaja v hrbtenjačo in v njej so skoncentrirani prehranjevalni, vazomotorični in dihalni centri.
Iz možganov odhaja 10 parov kranialnih živcev, kar je značilno za nižje vretenčarje.
Dvoživke
Dvoživke imajo številne progresivne spremembe v možganih, kar je povezano s prehodom na kopenski način življenja, kjer so razmere v primerjavi z vodnim okoljem bolj raznolike in za katere je značilna spremenljivost dejavnikov delovanja. To je vodilo do progresivnega razvoja čutil in s tem do progresivnega razvoja možganov.
^ Prednji možgani dvoživka je v primerjavi z ribami veliko večja, ima dve hemisferi in dva ventrikla. V strehi prednjih možganov so se pojavila živčna vlakna, ki tvorijo primarni medularni obok - arhipalij. Celična telesa nevronov se nahajajo globoko, obdajajo prekate, predvsem v striatumu. Vohalni režnji so še dobro razviti.
Najvišji integrativni center ostajajo srednji možgani (ihtiopsidni tip). Struktura je enaka kot pri ribah.
Mali možgani zaradi primitivnosti gibanja dvoživk ima obliko majhne plošče.
^ Intermediate in medulla oblongata enako kot pri ribah. Možgane zapušča 10 parov kranialnih živcev.
Plazilci (plazilci)
Plazilci pripadajo višjim vretenčarjem, zanje je značilen bolj aktiven življenjski slog, ki je povezan s postopnim razvojem vseh delov možganov.
^ Prednji možgani je največji del možganov. Pred njim segajo razviti vohalni režnji. Streha ostaja tanka, vendar se na medialni in lateralni strani vsake hemisfere pojavijo otoki korteksa. Lubje ima primitivno strukturo in se imenuje starodavno - arheokorteks. Vlogo višjega integrativnega centra opravljajo striatna telesa prednjih možganov - sauropsidnega tipa možgani. Striatum zagotavlja analizo dohodnih informacij in razvoj odzivov.
^ Vmesni, možgani, ker je povezan s pinealno žlezo in hipofizo, ima tudi hrbtni prirastek - parietalni organ, ki zaznava svetlobno stimulacijo.
Srednji možgani izgublja svoj pomen višjega integrativnega središča, zmanjšuje se tudi njegov pomen vizualnega središča, zato se zmanjšuje njegova velikost.
Mali možgani veliko bolje razvit kot pri dvoživkah.
^ Medulla oblongata tvori oster zavoj, značilen za višje vretenčarje, vključno s človekom.
Iz možganov odhaja 12 parov kranialnih živcev, kar je značilno za vse višje vretenčarje, vključno s človekom.
Ptice
Živčni sistem je zaradi splošne zapletenosti organizacije, prilagodljivosti na let in življenja v najrazličnejših okoljih veliko bolje razvit kot pri plazilcih.
Čez dan je za ptice značilno nadaljnje povečanje celotnega volumna možganov, zlasti prednjih možganov.
^ Prednji možgani pri ptice so najvišje integrativno središče. Njegov vodilni oddelek je striatum (sauropsidna vrsta možganov).
Streha ostaja slabo razvita. Ohranja le medialne otoke skorje, ki služijo kot najvišje vohalno središče. Potisnjeni so proti stičišču med poloblama in se imenujejo hipokampus. Vohalni režnji so slabo razviti.
^ diencefalon majhne velikosti in povezane s hipofizo in pinealno žlezo.
Srednji možgani ima dobro razvite optične režnje, kar je posledica vodilne vloge vida v življenju ptic.
Mali možgani velik, ima srednji del s prečnimi utori in majhnimi stranskimi izrastki.
^ Podolgovati Mot enako kot pri plazilcih. 12 parov kranialnih živcev.
Sesalci
Prednji možgani - to je največji del možganov. Pri različnih vrstah se njegove absolutne in relativne velikosti zelo razlikujejo. Glavna značilnost prednjih možganov je pomemben razvoj možganske skorje, ki zbira vse senzorične informacije iz čutil, proizvaja višjo analizo in sintezo teh informacij in postane aparat za subtilno pogojno refleksno aktivnost, pri visoko organiziranih sesalcih pa tudi za duševno aktivnost ( materinski tip možganov).
Pri najbolj organiziranih sesalcih ima skorja žlebove in vijuge, kar znatno poveča njeno površino.
D  Za prednje možgane sesalcev in ljudi je značilna funkcionalna asimetrija. Pri ljudeh se izraža v tem, da je desna hemisfera odgovorna za domišljijsko mišljenje, leva hemisfera pa za abstraktno mišljenje. Poleg tega se središča ustnega in pisnega govora nahajajo na levi hemisferi.
Za prednje možgane sesalcev in ljudi je značilna funkcionalna asimetrija. Pri ljudeh se izraža v tem, da je desna hemisfera odgovorna za domišljijsko mišljenje, leva hemisfera pa za abstraktno mišljenje. Poleg tega se središča ustnega in pisnega govora nahajajo na levi hemisferi.
^ diencefalon vsebuje približno 40 jeder. Posebna jedra talamusa obdelujejo vizualne, taktilne, okusne in interoceptivne signale, nato pa jih pošiljajo v ustrezna področja možganske skorje.
Hipotalamus vsebuje višje avtonomne centre, ki preko živčnih in humoralnih mehanizmov nadzirajo delovanje notranjih organov.
IN srednji možgani Kolikulus nadomesti kvadrigeminus. Njeni sprednji kolikuli so vidni, medtem ko so zadnji kolikuli povezani s slušnimi refleksi. V središču srednjih možganov je retikularna tvorba, ki služi kot vir naraščajočih vplivov, ki aktivirajo možgansko skorjo. Čeprav so sprednji režnji vizualni, se analiza vizualnih informacij izvaja v vidnih conah korteksa, srednji možgani pa v glavnem nadzorujejo očesne mišice - spremembe v lumnu zenice, gibanje oči in napetost akomodacije. V zadnjih gričih so centri, ki uravnavajo gibanje ušes, napetost bobniča in gibanje slušnih koščic. Srednji možgani sodelujejo tudi pri uravnavanju tonusa skeletnih mišic.
Mali možgani ima razvite stranske režnje (hemisfere), pokrite z lubjem, in črva. Mali možgani so povezani z vsemi deli živčnega sistema, ki so povezani z nadzorom gibanja - s prednjimi možgani, možganskim deblom in vestibularnim aparatom. Zagotavlja koordinacijo gibov.
^ Medulla oblongata . V njem so na straneh snopi živčnih vlaken, ki gredo v male možgane, na spodnji površini pa so podolgovati grebeni, imenovani piramide.
Obstaja 12 parov kranialnih živcev, ki izvirajo iz možganskega dna.
^ FILOGENEZA OBTOČIL
V večceličnih organizmih celice izgubijo neposreden stik z okoljem, zaradi česar je potreben sistem za prenos tekočine, ki celicam dostavi potrebne snovi in odstrani odpadne produkte. Pri nižjih nevretenčarjih (spužve, koelenterati, ploski črvi in okrogli črvi) poteka transport snovi z difuzijo tokov tkivne tekočine. Pri bolj organiziranih nevretenčarjih, pa tudi pri hordatih, se pojavijo posode, ki zagotavljajo kroženje snovi. Pojavi se krvožilni sistem, nato limfni sistem. Oba se razvijeta iz mezoderma.
Evolucijsko sta se razvili dve vrsti obtočil: zaprto in odprto. V zaprtem kri kroži samo skozi žile, v odprtem delu poti pa skozi režaste prostore - praznine in sinuse.
Krvožilni sistem se najprej pojavi pri kolobarjih. Zaprta je. Srca še ni. Obstajata dve glavni vzdolžni žili - trebušna in hrbtna, ki sta med seboj povezani z več obročastimi žilami, ki potekajo okoli črevesja. Manjše žile odstopajo od glavnih žil do organov, kri teče naprej skozi hrbtenično žilo in nazaj skozi trebušno žilo.
Pri členonožcih krvožilni sistem doseže višjo organizacijo. Imajo osrednji pulzirajoči aparat - srce, ki se nahaja na hrbtni strani telesa. Ko se skrči, vstopi kri v arterije, od koder se izliva v režaste prostore med organi (sinusi in praznine). nato se reabsorbira skozi parne odprtine v srcu, potem členonožci krvožilnega sistema niso zaprti.
Pri žuželkah kri ne opravlja funkcije transporta plinov, običajno je brezbarvna in se imenuje hemolimfa.
Tudi mehkužci imajo odprt krvni obtok, vendar imajo poleg arterij tudi venske žile. Srce ima več preddvorov, v katere se stekajo vene, in en velik prekat, iz katerega izhajajo arterije.
Pri najprimitivnejših hordatih, suličniku, krvožilni sistem v marsičem spominja na žilni sistem kolobarjev, kar kaže na njihovo filogenetsko povezanost. Lancelet nima srca, njegovo funkcijo opravlja trebušna aorta. Skozi to teče venska kri, ki vstopi v škržne žile, se obogati s kisikom in nato gre v hrbtno aorto, ki prenaša kri v vse organe. Venska kri iz sprednjega dela telesa se zbira v sprednjih, od zadnjega pa v zadnjih kardinalnih venah. Te vene se združijo v Cuvierjev kanal, skozi katerega kri vstopi v trebušno aorto.
V evoluciji vretenčarjev opazimo pojav srca, ki se nahaja na prsni strani telesa, in zaplet njegove strukture od dvokomorne do štirikomorne. Torej pri ribah je srce sestavljeno iz enega atrija in enega ventrikla, v njem teče venska kri. Obtok je samo en in kri se ne meša. Krvni obtok je v marsičem podoben krvožilnemu sistemu suličnika.
Pri kopenskih vretenčarjih se v povezavi s pridobivanjem pljučnega dihanja razvije drugi krog krvnega obtoka in srce poleg venske krvi začne prejemati arterijsko kri. V tem primeru se žilni sistem loči na obtočni in limfni.
Vmesno stopnjo v razvoju cirkulacijskega sistema od nižjih do višjih vretenčarjev zavzema cirkulacijski sistem dvoživk in plazilcev. Te živali imajo dva kroga krvnega obtoka, vendar v srcu pride do mešanja arterijske in venske krvi.
Popolna ločitev arterijske in venske krvi je značilna za ptice in sesalce, ki imajo štiriprekatno srce. Od dveh aortnih lokov, značilnih za dvoživke in plazilce, je ostal le eden: pri pticah je desni, pri sesalcih pa levi.
^ Razvoj arterijskih lokov.
Pri zarodkih vseh vretenčarjev se pred srcem oblikuje neparna trebušna aorta, iz katere izhajajo arterijski loki. So homologni arterijskim lokom suličnika. Toda njihovo število je manjše od števila lanceletov: pri ribah je 6-7 parov, pri kopenskih vretenčarjih pa 6 parov.
Prva dva para pri vseh vretenčarjih doživita redukcijo. Naslednji pari arterijskih lokov pri ribah so razdeljeni na aferentne in eferentne škržne arterije, pri kopenskih živalih pa so podvržene močnim transformacijam. Karotidne arterije torej nastanejo iz 3. para lokov. Četrti par je preoblikovan v aortne loke, ki se pri dvoživkah in plazilcih razvijajo simetrično. Pri pticah levi lok atrofira in se ohrani le desni. Pri sesalcih je desni lok reduciran in ohranjen le levi.
Peti par lokov je reduciran pri vseh vretenčarjih in le pri repatih dvoživkah ostane majhen kanal. Šesti lok izgubi povezavo s hrbtno aorto in iz njega izhajajo pljučne arterije. Posoda, ki med embrionalnim razvojem povezuje pljučno arterijo s hrbtno aorto imenovan botallov kanal. V odrasli dobi se ohrani pri repatih dvoživkah in nekaterih plazilcih. Kot razvojna napaka lahko ta kanal vztraja pri drugih bolj organiziranih živalih in ljudeh.
Limfni sistem je tesno povezan z obtočili: Limfa ima pomembno vlogo pri presnovi, saj je posrednik med krvjo in tkivno tekočino. Poleg tega je bogata z levkociti, ki igrajo pomembno vlogo pri imunosti.
^ RAZVOJ SRCA
V človeški embriogenezi opazimo številne filogenetske transformacije srca, kar je pomembno za razumevanje mehanizmov razvoja prirojenih srčnih napak.
Pri nižjih vretenčarjih (ribe, dvoživke) se srce nahaja pod žrelom v obliki votle cevi. Pri višjih vretenčarjih in človeku je srce oblikovano v obliki dveh cevi, ki sta daleč drug od drugega. Kasneje se približajo, se premikajo pod črevo, nato pa se zaprejo in tvorijo eno samo cev, ki se nahaja na sredini.
Pri vseh vretenčarjih iz sprednjega in zadnjega dela cevi nastanejo velike žile. Srednji del začne hitro in neenakomerno rasti in oblikuje S-obliko. Po tem se zadnji del cevi premakne na hrbtno stran in naprej ter tvori atrij. Sprednji del cevi se ne premika, njene stene se odebelijo in se spremeni v prekat.
Ribe imajo en atrij, pri dvoživkah pa je razdeljen na dvoje z rastočim septumom. Ribe in dvoživke imajo en prekat, vendar so v prekatu dvoživk mišični izrastki (trabekule), ki tvorijo majhne parietalne prekate. Pri plazilcih se v ventriklu oblikuje nepopolni septum, ki raste od spodaj navzgor.
Pri pticah in sesalcih je prekat razdeljen na dve polovici - desno in levo.
Med embriogenezo pri sesalcih in ljudeh sta na začetku en atrij in en ventrikel, ki sta drug od drugega ločena s prerezom s kanalom, ki povezuje atrij s prekatom. Nato začne v atriju od spredaj nazaj rasti septum, ki razdeli atrij na dva dela - levi in desni. Hkrati začnejo rasti izrastki na hrbtni in trebušni strani, ki sta povezani z dvema odprtinama: desno in levo. Kasneje se v teh luknjah oblikujejo ventili. Interventrikularni septum nastane iz različnih virov.
n  motnje srčne embriogeneze se lahko izrazijo v odsotnosti ali nepopolni fuziji interatrijskega ali interventrikularnega septuma. Od anomalij vaskularnega razvoja je najpogostejši odprti ductus botellus (od 6 do 22% vseh prirojenih malformacij kardiovaskularnega sistema), manj pogosto - odprti karotidni duktus. Poleg tega se lahko namesto enega aortnega loka razvijeta dva - levi in desni, ki tvorita aortni obroč okoli sapnika in požiralnika, s starostjo pa se lahko ta obroč zoži in požiranje je moteno. Včasih pride do transpozicije aorte, ko se ne začne iz levega prekata, ampak z desne, in pljučne arterije - z leve.
motnje srčne embriogeneze se lahko izrazijo v odsotnosti ali nepopolni fuziji interatrijskega ali interventrikularnega septuma. Od anomalij vaskularnega razvoja je najpogostejši odprti ductus botellus (od 6 do 22% vseh prirojenih malformacij kardiovaskularnega sistema), manj pogosto - odprti karotidni duktus. Poleg tega se lahko namesto enega aortnega loka razvijeta dva - levi in desni, ki tvorita aortni obroč okoli sapnika in požiralnika, s starostjo pa se lahko ta obroč zoži in požiranje je moteno. Včasih pride do transpozicije aorte, ko se ne začne iz levega prekata, ampak z desne, in pljučne arterije - z leve.
^ EVOLUCIJA ENDOKRINEGA SISTEMA
Usklajevanje dela organov in organskih sistemov pri živalih je zagotovljeno s prisotnostjo dveh tesno povezanih vrst regulacije - živčne in humoralne. Humoralno - je starejše in se izvaja skozi tekoče medije telesa s pomočjo biološko aktivnih snovi, ki jih izločajo celice in tkiva telesa v procesu presnove.
Z razvojem živali se je oblikoval poseben aparat za humoralni nadzor - endokrini sistem ali sistem endokrinih žlez. Od pojava slednjega živčna in humoralna regulacija delujeta v tesni medsebojni povezavi in tvorita en sam nevroendokrini sistem.
Hormonska regulacija, za razliko od živčne regulacije, je usmerjena predvsem v počasi potekajoče reakcije v telesu, zato ima vodilno vlogo pri regulaciji formativnih procesov: rasti, metabolizma, razmnoževanja in diferenciacije.
Pri nevretenčarjih se endokrine žleze najprej pojavijo v kolobarji. Najbolj raziskane endokrine žleze so pri rakih in žuželkah. Praviloma se endokrine žleze pri teh živalih nahajajo na sprednjem koncu telesa. U raki Obstajajo Y-organi, ki povzročajo taljenje. Te žleze so pod nadzorom X-organov, ki so funkcionalno tesno povezani z živčnimi gangliji glave. Poleg teh žlez imajo raki v očesnih pecljih sinusne žleze, ki uravnavajo procese preobrazbe.
U žuželke Na sprednjem koncu telesa so žleze z notranjim izločanjem, ki nadzorujejo preobrazbo in spodbujajo presnovo energije. Te žleze nadzira cefalična endokrina žleza, slednjo pa cefalični ganglij. Tako endokrini sistem rakov po svoji hierarhiji spominja na hipotalamo-hipofizni sistem vretenčarjev, kjer hipofiza uravnava delo vseh endokrinih žlez in je sama pod regulacijskim vplivom diencefalona.
E  endokrinih žlez vretenčarji
igrajo pomembnejšo vlogo pri regulaciji organskih sistemov kot pri nevretenčarjih. Poleg šestih ločenih endokrinih žlez (hipofiza, nadledvična žleza, ščitnica, obščitnične žleze, priželjc, epifiza) hormone proizvajajo številni organi, ki imajo še druge funkcije: spolne žleze, trebušna slinavka, nekatere celice prebavil, itd.
endokrinih žlez vretenčarji
igrajo pomembnejšo vlogo pri regulaciji organskih sistemov kot pri nevretenčarjih. Poleg šestih ločenih endokrinih žlez (hipofiza, nadledvična žleza, ščitnica, obščitnične žleze, priželjc, epifiza) hormone proizvajajo številni organi, ki imajo še druge funkcije: spolne žleze, trebušna slinavka, nekatere celice prebavil, itd.
Endokrine žleze pri vretenčarjih se v filogeniji razvijejo iz različnih virov in imajo različne lokacije. torej ščitnica žleza nastane iz epitelija ventralne strani žrela. Pri ribah se nahaja med prvo in drugo škržno režo, pri drugih vretenčarjih pa med drugo in tretjo škržno vrečko. Poleg tega je sprva ta žleza določena kot žleza z zunanjim izločanjem. Med filogenezo pri številnih vretenčarjih ščitnična žleza spremeni svojo lokacijo in, začenši z dvoživkami, se v njej pojavijo režnji in ožina, kar ni značilno za ribe, kjer je videti kot ena vrvica.
^ Thymus žleza (timus) pri ribah se razvije zaradi epitelijskih izrastkov, ki nastanejo na stenah vseh škržnih vrečk. Ti izrastki se kasneje zavežejo in tvorijo dva ozka trakova, sestavljena iz limfoidnega tkiva, z lumnom v notranjosti.
Pri dvoživkah in plazilcih je število zametkov, iz katerih se razvije timus, znatno zmanjšano - izvirajo iz drugega in tretjega para škržnih vrečk. Pri sesalcih - iz treh parov škržnih vrečk, vendar predvsem iz drugega para.
hipofiza pri kopenskih vretenčarjih je sestavljen iz treh režnjev: sprednjega, srednjega (vmesnega) in zadnjega; in v ribah - samo od spredaj in od sredine.
Hipofiza je povezana s spodnjo površino diencefalona in se razvije iz različnih virov, sprednji in srednji reženj iz epitelija strehe ustne votline, zadnji reženj pa iz distalnega dela infundibuluma diencefalona (nevralni izvor). Funkcija hipofize pri ribah je samo proizvodnja gonadotropnih hormonov (spodbujanje proizvodnje spolnih hormonov v spolnih žlezah). Dvoživke razvijejo zadnji reženj, kar je razloženo z njihovim prehodom na kopenski način življenja in potrebo po uravnavanju metabolizma vode. Aksoni nevrosekretornih nevronov hipotalamusa vstopijo v posteriorni reženj in antidiuretični hormon, ki ga izločajo, se kopiči in nato vstopi v kri.
Srednji reženj, ki se začne pri dvoživkah, izgubi sposobnost proizvodnje gonadotropnega hormona in zdaj proizvaja hormon, ki spodbuja sintezo melanina. Pri kopenskih vretenčarjih sprednji reženj poleg gonadotropina izloča še druge tropske hormone, pa tudi rastni hormon.
Nadledvične žleze pri Hordati se razvijejo iz dveh virov. Njihovo skorjo tvori epitelij peritoneja, medula pa je nevralnega izvora. Poleg tega se pri ribah kortikalna snov nahaja vzdolž hrbtne površine primarnih ledvic metamerično in ločeno druga od druge, medula pa se nahaja v bližini genitalnih grebenov na obeh straneh mezenterija.
Pri dvoživkah nastane prostorska povezava med telesi nadledvične žleze, pri amniotih pa se združijo vsi anlagi nadledvične žleze in tvorijo parni organ, sestavljen iz zunanje skorje in notranje medule. Nadledvične žleze se nahajajo nad zgornjim polom ledvic.
^ EVOLUCIJA IMUNSKEGA SISTEMA
Imunski sistem ščiti telo pred prodiranjem genetsko tujih teles: mikroorganizmov, tujih celic, tujkov itd. Njegovo delovanje temelji na sposobnosti razlikovanja lastnih telesnih struktur od genetsko tujih in slednje odpravlja.
V evoluciji so se oblikovale tri glavne oblike imunskega odziva: 1) fagocitoza ali nespecifično uničenje gensko tujega materiala; 2) celična imunost, ki temelji na njenem specifičnem prepoznavanju in uničenju s strani limfocitov T; 3) humoralna imunost, ki se izvaja s pretvorbo B-limfocitov v plazemske celice in njihovo sintezo protiteles (imunoglobulinov).
V evoluciji obstajajo tri stopnje oblikovanja imunskega odziva:
- kvaziimuno (latinsko "quasi" - podobno) priznanje lastne in tuje celice telesa. Ta vrsta reakcije je opažena od coelenteratov do sesalcev. S tem odzivom se ne oblikuje imunski spomin, to pomeni, da ni krepitve imunskega odziva na ponavljajoče se prodiranje tujega materiala;
p primitivna celična imunost najdemo v anelidah in iglokožcih. Zagotavljajo ga kolomociti - celice sekundarne telesne votline, ki so sposobne uničiti tujek. Na tej stopnji se pojavi imunološki spomin;
sistem integrirane celične in humoralne imunosti. Zanjo so značilne specifične humoralne in celične reakcije na tujke, prisotnost limfoidnih imunskih organov in tvorba protiteles. Ta vrsta imunskega sistema ni značilna za nevretenčarje.
Timus, vranica in posamezne akumulacije limfoidnega tkiva najdemo v celoti, začenši z dvoživkami. Pri nižjih vretenčarjih (ribe, dvoživke) žleza timus aktivno izloča protitelesa, kar ni značilno za ptice in sesalce.
Posebnost imunskega sistema imunskega odziva ptic je prisotnost posebnega limfoidnega organa - Fabriciusove burze. V tem organu se lahko limfociti B po antigenski stimulaciji spremenijo v plazmatke, ki proizvajajo protitelesa.
Pri sesalcih so organi imunskega sistema razdeljeni na 2 tipa: centralni in periferni. V osrednjih organih imunogeneze se zorenje limfocitov pojavi brez vpliva antigenov. V perifernih organih imunogeneze pride do antigena odvisnih T in B - razmnoževanje in diferenciacija limfocitov.
V zgodnjih fazah embriogeneze limfne matične celice migrirajo iz rumenjakove vrečke v timus in rdeči kostni mozeg. Po rojstvu je vir izvornih celic rdeči kostni mozeg. Periferni limfoidni organi so: bezgavke, vranica, mandlji, črevesni limfoidni mešički. Ob rojstvu so še praktično neoblikovane in razmnoževanje in diferenciacija limfocitov v njih se začne šele po antigenski stimulaciji T- in B-limfocitov, ki so se preselili iz osrednjih organov imunogeneze.
^ RAZVOJ DIHALNEGA SISTEMA .
Skoraj vsi živi organizmi so aerobi, to je dihalci zraka. Skupek procesov, ki zagotavljajo vnos in porabo O 2 ter sproščanje CO 2, imenujemo dihanje.
Dihalna funkcija je različno zagotovljena pri živalih različnih stopenj organiziranosti. Najenostavnejša oblika dihanja je difuzija plinov skozi stene žive celice (pri enoceličnih organizmih) ali skozi ovojnice telesa (kolenterati; ploščati, okrogli in obročasti črvi). Difuzno dihanje najdemo tudi pri majhnih členonožcih, ki imajo tanek hitinski pokrov in razmeroma veliko telesno površino.
Ker postaja organizacija živali bolj zapletena, se oblikuje poseben dihalni sistem; Tako se že v nekaterih vodnih kolobarjih pojavijo primitivni dihalni organi - zunanje škrge (epitelijski izrastki s kapilarami), pri dihanju pa sodeluje tudi koža. Pri členonožcih imajo dihala bolj zapleteno zgradbo in jih pri vodnih oblikah predstavljajo škrge, pri kopenskih in sekundarnih vodnih oblikah pa pljuča in sapnik (pri najstarejših členonožcih, kot so škorpijoni, pljuča, pri pajkih obe pljuča oz. sapnik, pri žuželkah pa višji členonožci - samo sapnik).
Funkcijo dihalnih organov pri nižjih strunastih (suličnikih) prevzamejo škržne reže, po katerih pregradah potekajo škržne arterije (100 parov). Ker v škržnih pregradah ni delitve arterij na kapilare, je skupna površina za vnos O2 majhna in oksidativni procesi potekajo na nizki ravni. V skladu s tem lancelet vodi sedeč življenjski slog.
V zvezi s prehodom vretenčarji Za Z aktivnim življenjskim slogom pride do progresivnih sprememb v dihalnih organih. Torej, v ribah v škržnih filamentih se v nasprotju z lanceletom pojavi obilna mreža krvnih kapilar, njihova dihalna površina se močno poveča, zato se število škržnih rež pri ribah zmanjša na štiri.
Dvoživke - prve živali, ki so prišle na kopno, ki so razvile atmosferske dihalne organe - pljuča (iz izrastka črevesne cevi). Zaradi primitivnosti zgradbe (pljuča so vrečke s tankimi celičnimi stenami) količina kisika, ki vstopa skozi pljuča, zadovolji potrebe telesa po njem le za 30-40 %, zato koža, ki vsebuje številne krvne kapilare (dermalne kapilare), sodeluje tudi pri pljučnem dihanju).
Dihalne poti dvoživk so slabo diferencirane. Z orofarinksom so povezani z majhno laringealno-trahealno komoro.
^
U  plazilci
v povezavi s končnim izstopom na kopno se dihalni sistem še bolj zaplete: kožno dihanje izgine, dihalna površina pljučnih vrečk se poveča zaradi pojava velikega števila razvejanih predelov, v katerih prehajajo krvne kapilare. Tudi dihalne poti postanejo bolj zapletene: v sapniku se oblikujejo hrustančni obročki, ki se delijo, nastaneta dva bronhija. Začne se nastajanje intrapulmonalnih bronhijev.
plazilci
v povezavi s končnim izstopom na kopno se dihalni sistem še bolj zaplete: kožno dihanje izgine, dihalna površina pljučnih vrečk se poveča zaradi pojava velikega števila razvejanih predelov, v katerih prehajajo krvne kapilare. Tudi dihalne poti postanejo bolj zapletene: v sapniku se oblikujejo hrustančni obročki, ki se delijo, nastaneta dva bronhija. Začne se nastajanje intrapulmonalnih bronhijev.
U  ptice
V strukturi dihalnih organov se pojavijo številne značilnosti. Njihova pljuča imajo številne pregrade z mrežo krvnih kapilar. Iz sapnika izhaja bronhialno drevo, ki se konča z bronhiolami. Del glavnih in sekundarnih bronhijev sega čez pljuča in tvori cervikalne, torakalne in trebušne pare zračnih vrečk ter prodira tudi v kosti in jih naredi pnevmatične. Med letom je kri nasičena s kisikom tako pri vdihu kot pri izdihu (dvojno dihanje).
ptice
V strukturi dihalnih organov se pojavijo številne značilnosti. Njihova pljuča imajo številne pregrade z mrežo krvnih kapilar. Iz sapnika izhaja bronhialno drevo, ki se konča z bronhiolami. Del glavnih in sekundarnih bronhijev sega čez pljuča in tvori cervikalne, torakalne in trebušne pare zračnih vrečk ter prodira tudi v kosti in jih naredi pnevmatične. Med letom je kri nasičena s kisikom tako pri vdihu kot pri izdihu (dvojno dihanje).
Sesalci imajo pljuča z alveolarno zgradbo, zaradi česar je njihova površina 50-100-krat večja od površine telesa. Bronhiji so drevesasto razvejani in se končajo s tankostenskimi bronhiolami z gručami alveolov, gosto prepletenih s krvnimi kapilarami. Grlo in sapnik sta dobro razvita.
Tako je glavna smer razvoja dihalnega sistema povečanje dihalne površine, zapletanje strukture dihalnih poti in njihova ločitev od dihalnih.
^ RAZVOJ IZLOČALNEGA SISTEMA
Pri enoceličnih živalih in coelenteratah procesi sproščanja strupenih presnovnih produktov potekajo z difuzijo iz celic v zunajcelično okolje. Vendar pa se že pri ploščatih črvih pojavi sistem tubulov, ki opravljajo izločevalne in osmoregulacijske funkcije. Ti tubuli se imenujejo protonefridij. Začnejo se z veliko zvezdasto celico, v citoplazmi katere je tubul s šopom migetalk, ki ustvarjajo tok tekočine. Te celice izvajajo aktivni transport in osmozo vode in raztopljenih škodljivih snovi v lumen citoplazemskega tubula.
Izločevalni sistem okrogli črvi imajo tudi protonefridijski značaj.
Pri anelidah so organi izločanja in osmoregulacije metanefridija. To so tubuli, katerih en konec je razširjen v obliki lijaka, obdan z migetalkami in obrnjen proti telesni votlini, drugi konec pa se odpira na površini telesa z izločevalno poro. Tekočina, ki jo izločajo tubuli, se imenuje urin. Nastane s filtracijo – selektivno reabsorpcijo in aktivnim izločanjem iz tekočine v telesni votlini. Metanefridialni tip izločevalnega sistema je značilen tudi za ledvice mehkužcev.
Pri členonožcih so izločevalni organi bodisi modificirani metanefridiji oz Malpigijci plovila, oz specializirane žleze
Malpigijeve žile so snop cevk, katerih en konec se slepo konča v telesni votlini in vsrkava izločane produkte, drugi konec pa se odpre v črevesno cev.
Razvoj izločevalnega sistema hordatov se izraža v prehodu iz nefridijev nižjih hordatov v posebne organe - ledvice.
Suličnik ima izločevalni sistem, podoben izločevalnemu sistemu kolobarjev. Predstavljena je s 100 pari nefridij, katerega en konec je obrnjen proti sekundarni telesni votlini in absorbira izločevalne produkte, drugi pa jih odvaja v peribranhialno votlino.
Izločevalni organi vretenčarjev so seznanjeni popki. Pri nižjih vretenčarjih (ribe, dvoživke) se med embriogenezo tvorita dve vrsti ledvic: prednost (ali glava ledvice) in trup (ali primarni). Prednost je po strukturi podobna metanefridiji. Sestavljen je iz zavitih tubulov, ki so usmerjeni v telesno votlino, drugi konec pa se izliva v skupni kanal ledvice. Nedaleč od vsakega lijaka je vaskularni glomerul, ki filtrira presnovne produkte v telesno votlino. Ta vrsta ledvice deluje le v obdobju ličinke, nato pa začne delovati primarna ledvica. V njem so vzdolž ledvičnih tubulov izbokline, v katerih se nahajajo žilni glomeruli in se filtrira urin. Lijaki izgubljajo funkcionalni pomen in se zaraščajo.
Pri višjih vretenčarjih se v embrionalnem obdobju zaporedno oblikujejo tri ledvice: prednost, primarni (torzo) in sekundarna (medenična) ledvica. Ledvica ne deluje. Primarna ledvica deluje le med embriogenezo. Njen kanal se razcepi na dva: Wolffov in Müllerjev kanal. Nato se Wolffovi kanali spremenijo v ureterje in pri moških v sečevode in vas deferens. Müllerjevi kanali so ohranjeni le pri samicah in se preoblikujejo v jajcevode. Tisti. Med embriogenezo sta urogenitalni in reproduktivni sistem povezana.
Proti koncu embrionalnega obdobja začne delovati medenična (sekundarna) ledvica. To so kompaktne seznanjene formacije, ki se nahajajo na straneh ledvene hrbtenice. Morfo-funkcionalna enota v njih je nefron, ki je sestavljen iz kapsule z vaskularnim glomerulom sistema zvitih tubulov prvega in drugega reda in Henlejeve zanke. Tubuli nefrona postanejo zbiralni kanali, ki se odpirajo v ledvično medenico.
^ EVOLUCIJA IMUNSKEGA SISTEMA
Imunski sistem ščiti telo pred prodiranjem genetskih tujkov v telo: mikroorganizmi, virusi, tujki, tujki. Njegovo delovanje temelji na sposobnosti razlikovanja lastnih struktur od genetsko tujih in jih odpravlja.
V evoluciji so se pojavile tri glavne oblike imunskega odziva:
fagocitoza – ali nespecifično uničenje genetsko tujega
celična imunost, ki temelji na specifičnem prepoznavanju in uničenju takšnega materiala s strani limfocitov T;
humoralna imunost, ki se izvaja s tvorbo potomcev B-limfocitov, tako imenovanih plazemskih celic imunoglobulinov in njihove vezave tujih antigenov.
Faza I - kvaziimunsko (latinsko quasi - kot, kot da) prepoznavanje lastnih in tujih celic s strani telesa. Ta vrsta reakcije je opažena od coelenteratov do sesalcev. Ta reakcija ni povezana s proizvodnjo imunskih teles in imunski spomin se ne oblikuje, to pomeni, da ni krepitve imunskega odziva na ponavljajoče se prodiranje tujega materiala.
Stopnja II - primitivna celična imunost najdemo pri anelidih in iglokožcih. Zagotavljajo ga kolomociti - celice sekundarne telesne votline, ki so sposobne uničiti tujek. Na tej stopnji se pojavi imunološki spomin.
Faza III - sistem integrirane celične in humoralne imunosti. Zanj so značilne specifične humoralne in celične reakcije na tujke. Zanj je značilna prisotnost limfoidnih imunskih organov in tvorba protiteles. Ta vrsta imunskega sistema ni značilna za nevretenčarje.
Ciklostomi so sposobni tvoriti protitelesa, vendar je še vedno odprto vprašanje, ali imajo priželjc kot osrednji organ imunogeneze. Timus so najprej odkrili pri ribah.
Evolucijske predhodnike limfoidnih organov sesalcev - timusa, vranice, kopičenja limfoidnega tkiva najdemo v celoti v dvoživke. Pri nižjih vretenčarjih (ribe, dvoživke) žleza timus aktivno izloča protitelesa, kar ni značilno za ptice in sesalce.
Značilnosti imunskega odzivnega sistema ptice je sestavljen iz prisotnosti posebnega limfoidnega organa - Fabrician torba. Ta organ proizvaja limfocite B, ki se po antigenski stimulaciji lahko spremenijo v plazmatke in tvorijo protitelesa.
U sesalci Organi imunskega sistema so razdeljeni na dve vrsti: centralni in periferni. V osrednjih organih poteka zorenje limfocitov brez pomembnega vpliva antigenov. Razvoj perifernih organov je, nasprotno, neposredno odvisen od antigenskega učinka - šele ob stiku z antigenom se v njih začnejo procesi proliferacije in diferenciacije limfocitov.
Osrednji organ imunogeneze pri sesalcih je timus, kjer nastajajo limfociti T, ter rdeči kostni mozeg, kjer nastajajo limfociti B.
V zgodnjih fazah embriogeneze limfne matične celice migrirajo iz rumenjakove vrečke v timus in rdeči kostni mozeg. Po rojstvu je vir izvornih celic rdeči kostni mozeg.
Periferni limfoidni organi so: bezgavke, vranica, mandlji, črevesni limfoidni mešički. Ob rojstvu še niso praktično oblikovani in se v njih začne nastajanje limfocitov šele po antigenski stimulaciji, potem ko se naselijo z limfociti T in B iz osrednjih organov imunogeneze.
^ FILOGENEZA VISCERIALNE LOBANJE PRI VRETENČARJIH .
Lobanja vretenčarjev je sestavljena iz dveh glavnih delov - aksialnega in visceralnega.
Aksialni - kranij (možganska lobanja - nevrokranij) - nadaljevanje aksialnega skeleta, služi za zaščito možganov in čutnih organov.
Visceralno-obrazni (splanhnokranij), tvori oporo za sprednji del prebavnega trakta.
 ba deli lobanje se razvijajo neodvisno drug od drugega, na različne načine. Visceralni del lobanje pri zarodkih vretenčarjev je sestavljen iz metamerno nameščenih hrustančnih lokov, ki pokrivajo sprednji del prebavnega trakta in so med seboj ločeni z visceralnimi režami. Loki so označeni z zaporednimi številkami glede na njihovo lokacijo glede na lobanjo.
ba deli lobanje se razvijajo neodvisno drug od drugega, na različne načine. Visceralni del lobanje pri zarodkih vretenčarjev je sestavljen iz metamerno nameščenih hrustančnih lokov, ki pokrivajo sprednji del prebavnega trakta in so med seboj ločeni z visceralnimi režami. Loki so označeni z zaporednimi številkami glede na njihovo lokacijo glede na lobanjo. Prvi lok pri večini sodobnih vretenčarjev dobi funkcijo čeljustnega aparata - imenuje se maksilarni, drugi pa - tudi glede na njegovo funkcijo - hioid ali hioid. Ostali, od tretjega do sedmega, se imenujejo škrge, ker. služijo kot podpora za škržni aparat. V zgodnjih fazah razvoja visceralna in aksialna lobanja nista povezani drug z drugim;
Zarodek sedmih visceralnih lokov med embrionalnim razvojem je skupen vsem zarodkom vretenčarjev pri predstavnikih različnih razredov.
^I. Slabše ribe (hrustančne) - Chondrichthyes
1., znan tudi kot čeljustni lok, je sestavljen iz dveh velikih hrustancev, podolgovatih v anteroposteriorni smeri: zgornji - palatokvadrat - primarna zgornja čeljust, spodnji - Meckelov - primarna spodnja čeljust; zadaj so med seboj zraščene in opravljajo funkcijo primarne čeljusti.
2., znan tudi kot hioid ali hioidni lok, je sestavljen iz naslednjih komponent:
dveh, ki se nahajata na vrhu hiomandibularnega hrustanca, ki sta od zgoraj povezana z lobanjo, spodaj - s hioidom in spredaj - s čeljustnim lokom - primarna zgornja čeljust;
dveh hioidov, ki se nahajajo pod hiomandibularnimi hrustanci, ki so povezani z njimi; poleg tega so hioidi povezani s primarno spodnjo čeljustjo;
iz neparne kopule (majhen hrustanec, ki povezuje obe podjeznici med seboj).
Preostali visceralni loki od tretjega do sedmega tvorijo oporo dihalnemu aparatu.
^ II Višje ribe - (koščene) Osteichthyes .
Glavna razlika zadeva samo čeljustni lok:
1) zgornji element čeljustnega loka (zgornja čeljust) je namesto enega velikega palatinskega kvadratnega hrustanca sestavljen iz petih elementov - palatinskega hrustanca, kvadratne kosti in treh pterigoidnih hrustancev;
pred primarno zgornjo čeljustjo nastaneta dve veliki zgornji kosti, opremljeni z velikimi zobmi - te kosti postanejo sekundarne zgornje čeljusti;
distalni konec primarne spodnje čeljusti prav tako pokriva velika zobna kost, ki štrli daleč naprej in tvori sekundarno spodnjo čeljust. Hioidni lok ohranja svojo prejšnjo funkcijo, tj. lobanja ostane hipostilna.
Glavna razlika je v novem načinu povezave čeljustnega loka z lobanjo: nebni hrustanec primarne zgornje čeljusti se po vsej dolžini zlije z aksialno lobanjo, tj. z lobanjo. Ta vrsta povezave se imenuje avtoslog.
Mandibularni odsek je povezan z maksilarnim odsekom in ima tudi povezavo z lobanjo brez hioidnega loka.
Tako se hiomandibularni hrustanec osvobodi funkcije obešanja, se znatno zmanjša in pridobi novo funkcijo - je del zračne votline srednjega ušesa v obliki slušne koščice - kolone.
Del hioidnega loka (hioidni hrustanec), vejni loki tvorijo delno oporo za jezik in hioidni aparat, delno so laringealni hrustanci delno reducirani.
^ IV. Plazilci - Reptilia .
Lobanja je avtostilna, hkrati pa je nebni hrustanec primarne čeljusti reduciran in pri artikulaciji zgornje čeljusti z lobanjo sodeluje samo kvadratna kost, spodnja čeljust pa je povezana z njo in se tako združuje z lobanjo. Preostali del visceralnega okostja tvori hioidni aparat, ki ga sestavljajo telo hioidne kosti in trije pari procesov.
^ V. Sesalci - Mammalia .
Pojavi se popolnoma nov način povezovanja z lobanjo spodnje čeljusti, ki se pritrdi neposredno nanjo in tvori sklep s skvamozalno kostjo lobanje, kar omogoča ne le prijemanje hrane, temveč tudi izvajanje kompleksnih žvečilnih gibov. Pri nastanku sklepa sodeluje samo sekundarna spodnja čeljust. Posledično kvadratna kost primarne maksile izgubi funkcijo obešanja in se spremeni v slušno koščico - inkus.
Med embrionalnim razvojem primarna spodnja čeljust popolnoma zapusti spodnjo čeljust in se preoblikuje v naslednjo slušno kost - malleus.
Zgornji del hioidnega loka, homologa hiomandibularnega hrustanca, se spremeni v streme.
Vse tri slušne koščice tvorijo eno samo funkcionalno verigo.
1. - vejni lok (1. visceralni) in kopula povzročita nastanek telesa hioidne kosti in njenih zadnjih rogov.
Iz 2. in 3. kračnega loka (4. in 5. visceralni) nastane ščitasti hrustanec, ki se najprej pojavi pri sesalcih.
4. in 5. vejni lok (1. in 7. visceralni) zagotavljata material za preostale laringealne hrustance in po možnosti za trahealne hrustance.
^ EVOLUCIJA ZOBNEGA SISTEMA
IN USTNE ŽLEZE VRETENČARJEV
Ribi-Ribi
Zobni sistem je homodonten (zobje so enaki). Zobje so stožčaste oblike, obrnjeni nazaj, služijo zadrževanju hrane, se nahajajo vzdolž roba lobanje, v nekaterih primerih pa na celotni površini ustne votline.
V ustni votlini ni žlez slinavk, ker... Hrano pogoltnejo z vodo. Jezik je primitiven, v obliki dvojne gube sluznice. Streho ustne votline tvori lobanjska osnova – primarno trdo nebo. Ustno odprtino obdajajo kožne gube – ustnice, ki so negibne. Splošna orofaringealna votlina.
Plakoidne luske hrustančnic so plošča, na kateri je nameščena bodica. Plošča leži v koriumu, vrh hrbtenice štrli skozi povrhnjico. Celotna luska je sestavljena iz dentina, ki ga tvorijo celice koriuma; konica bodice je prekrita s sklenino, ki jo tvorijo celice bazalne plasti povrhnjice.
Večje in bolj zapletene plakoidne luske se nahajajo v čeljusti in tvorijo zobe. V bistvu so zobje vseh vretenčarjev spremenjene plakoidne luske njihovih prednikov.
Dvoživke - Dvoživke.
Zobni sistem homodont. Zobje številnih dvoživk se nahajajo ne le na alveolarnem loku; polifiodontija.
Pojavijo se žleze slinavke, katerih izloček ne vsebuje encimov. Jezik vsebuje mišice, ki določajo njegovo lastno gibljivost. Strop ustne votline je tudi primarno trdo nebo. Splošna orofaringealna votlina je nepremična.
Plazilci- Plazilci.
Zobni sistem pri sodobnih plazilcih homodont, Strupeni plazilci imajo posebne zobe, skozi katere strup teče v ugrizno rano. Zobje so razporejeni v eni vrsti. Nekatere izumrle oblike kažejo začetno diferenciacijo. Značilen za vse plazilce polifiodontija.
Žleze slinavke so bolje razvite, med njimi so podjezične, zobne in labialne. Izloček žlez že vsebuje encime.
Pri strupenih kačah se zadnji par zobnih žlez spremeni v strupene, izloček vsebuje toksine (strup).
Jezik je sestavljen iz treh rudimentov: enega - neparnega in dveh - seznanjenih, ki ležita pred neparnim. Parni primordiji se kasneje zrastejo. Pri večini plazilcev je to zlitje nepopolno in jezik je razcepljen.
Zametki sekundarnega trdega neba se pojavijo v obliki vodoravnih kostnih gub zgornje čeljusti, ki segajo do sredine in delijo ustno votlino na zgornji del - dihalni (nazofaringealni) in spodnji - sekundarno ustno votlino. Ustnice so nepremične.
Sesalci- sesalec,
Zobje heterodont, tiste. diferencirani: obstajajo sekalci (incisivi), očesci (canini), mali kočniki (praemolares) in kočniki (molares). Pri plavutonožcih in zobatih kitih se zobje ne razlikujejo. Zobje sedijo v alveolnih lokih čeljusti, osnova zoba se zoži in tvori korenino.
Sekalci in očesci so zelo podobni stožčastim zobem svojih prednikov (plazilcev), kočniki so doživeli največje evolucijske preobrazbe in so se prvič pojavili pri kuščarjih z zobmi.
IN  Zaradi diferenciacije zob se trajanje delovanja poveča. V ontogenezi pride do dveh menjav zob ( difiodontija): sekalci, očesci in veliki kočniki imajo dve generaciji (listopadno in stalno); majhni radikali - samo eden.
Zaradi diferenciacije zob se trajanje delovanja poveča. V ontogenezi pride do dveh menjav zob ( difiodontija): sekalci, očesci in veliki kočniki imajo dve generaciji (listopadno in stalno); majhni radikali - samo eden.
Skupno število zob se razlikuje med različnimi vrstami: na primer, sloni jih imajo 6, volkovi 42, mačke 30, zajci 28, večina primatov in ljudi jih ima 32.
Žleze slinavke sesalcev so številne: to so majhne - lingvalne, bukalne, palatalne, zobne - homologne žlezam plazilcev, in velike - sublingvalne, submandibularne, parotidne. Od teh sta se prvi dve pojavili kot posledica diferenciacije sublingvalne žleze plazilcev, parotidne žleze pa so bile nova pridobitev sesalcev. V ustni votlini - pri višjih sesalcih se pojavijo velike kopice limfnega tkiva - tonzile.
Jezik se tako kot pri plazilcih razvije iz treh začetkov. Sekundarno trdo nebo postane neprekinjeno, ustna votlina je popolnoma ločena od nosne votline, s čimer dosežemo neodvisnost funkcij ustne votline in dihanja. Zadaj se trdo nebo nadaljuje v mehko nebo – dvojno gubo sluznice, ki ločuje ustno votlino od žrela. Prečni grebeni trdega neba prispevajo k mletju hrane. Pri ljudeh po rojstvu postopoma izginejo.
Ustnice vrečarjev in posteljic so mesnate in mobilne, kar je povezano s hranjenjem mladičev z mlekom. Ustnice, lica in čeljusti določajo prostor, imenovan preddverje ust.
^ Pri ljudeh zobna formula 2123
2123(polovica zgornje in spodnje čeljusti).
Zobje so se v primerjavi z drugimi primati zmanjšali, zlasti očesci ne štrlijo iz zobovja in se ne prekrivajo. Diasteme (razmiki med zobmi) v zgornji in spodnji čeljusti so izginile, zobje so postali v gosti vrsti, zobni lok je dobil zaobljeno (parabolično) obliko.
M  polare imajo obliko štirih gomoljev. Zadnji par kočnikov, "modrostnih zob", izbruhne pozno - do 25 let. So očitno rudimentarni, zmanjšani in pogosto slabo diferencirani.
polare imajo obliko štirih gomoljev. Zadnji par kočnikov, "modrostnih zob", izbruhne pozno - do 25 let. So očitno rudimentarni, zmanjšani in pogosto slabo diferencirani.
Med žvečenjem lahko spodnja čeljust izvaja rotacijske gibe glede na zgornjo zaradi neprekrivanja zmanjšanih očescev in komplementarnih konic žvečilnih zob obeh čeljusti.
^ ATAVISTIČNE ANOMALIJE ČLOVEŠKE USTNE VOTLINE:
A) redka anomalija - homodontni zobni sistem, vsi zobje so stožčasti;
B) trikuspidalni kočniki;
C) izraščanje presežnih zob, tj. pri človeku je možno oblikovati več kot 32 zobnih kalčkov;
D) odsotnost "modrostnih zob";
D) zelo redka malformacija jezika - bifurkacija njegovega konca, ki je posledica nezlitja parnih rudimentov v embriogenezi;
E) motnje zlitja (to bi se moralo zgoditi do konca osmega tedna embriogeneze) vodoravnih kostnih gub, ki tvorijo trdo nebo, kar vodi v nezalitje trdega neba in nastanek okvare, znane kot "razcep" okus«;
G) razcep zgornje ustnice (»razcep ustnice«) nastane zaradi nepopolnega zlitja dermalno-mezodermalnih procesov, ki tvorijo zgornjo ustnico, od katerih dva (bočna) rasteta iz zgornje čeljusti, eden (osrednji) pa iz čeljusti. frontonazalni proces.
^ SINTETIČNA TEORIJA EVOLUCIJE
Kombinacija darvinizma z ekologijo in genetiko, ki se je začela v dvajsetih letih prejšnjega stoletja, je utrla pot nastanku sintetične teorije evolucije, ki je danes edina celostna, dokaj v celoti razvita teorija biološke evolucije, ki uteleša klasični darvinizem in populacijsko genetiko.
Prvi znanstvenik, ki je uvedel genetski pristop k preučevanju evolucijskih procesov, je bil Sergej Sergejevič Četverikov. Leta 1926 je objavil znanstveni članek »O nekaterih vidikih evolucijskega procesa z vidika sodobne genetike«, v katerem je na primeru naravnih populacij Drosophila pokazal, da: 1) se nenehno pojavljajo mutacije v naravnih populacijah; 2) recesivne mutacije vrste "vpijejo kot goba" in v heterozigotnem stanju lahko vztrajajo za nedoločen čas; 3) ko se vrsta stara, se v njej kopiči vedno več mutacij in značilnosti vrste se rahljajo; 4) izolacija in dedna variabilnost sta glavna dejavnika intraspecifične diferenciacije; 5) panmiksija vodi v polimorfizem vrste, selekcija pa v monomorfizem. V tem delu S. S. Chetverikov poudarja, da kopičenje majhnih naključnih mutacij s selekcijo vodi do naravnega, prilagodljivo usmerjenega poteka evolucije. Dela S.S. Chetverikova so nadaljevali domači genetiki, kot je N.V. Timofejev-Resovski, D.D. Romashov. N.P. Dubinin, N.I. Vavilov in drugi. Ta dela so utrla pot za ustvarjanje temeljev sintetične teorije evolucije.
V 30. letih so dela angleških znanstvenikov R. Fisherja. J. Holdame. S. Wright je postavil temelje za sintezo teorije evolucije in genetike na Zahodu.
Eno prvih del, ki je orisalo bistvo sintetične teorije evolucije, je bila monografija F. G. Dobzhanskega "Genetika in izvor vrst" (1937). struktura populacij glede na vpliv takšnih dejavnikov in vzrokov evolucije, kot so dedna variabilnost, naravna selekcija, nihanje števila osebkov v populacijah (populacijski valovi), migracije in končno reproduktivna izolacija novih oblik, ki so nastale znotraj vrste.
Izjemen prispevek k ustvarjanju sintetične teorije evolucije je prispeval domači znanstvenik I.I. Schmalhausen. Na podlagi ustvarjalne kombinacije evolucijske teorije, embriologije, morfologije, paleontologije in genetike je poglobljeno raziskal odnos med ontogenezo in filogenijo, preučeval glavne smeri evolucijskega procesa in razvil številne temeljne določbe sodobne teorije evolucije. Njegova glavna dela: "Organizem kot celota v individualnem in zgodovinskem razvoju" (1938); "Poti in vzorci evolucijskega procesa" (1939); "Dejavniki evolucije" (1946).
Pomembno mesto med temeljnimi študijami o teoriji evolucije zavzema monografija "Evolution Modern Synthesis" (1942), ki je izšla leta 1942 pod vodstvom uglednega angleškega evolucionista Juliana Huxleyja, pa tudi študije o stopnjah in oblikah. evolucije, ki se jo je lotil George Simpson,
Sintetična teorija evolucije temelji na 11 osnovnih postulatih, ki jih je v jedrnati obliki oblikoval sodobni domači genetik N.N. Vorontsov, približno v tej obliki:
1. Gradivo za evolucijo so praviloma zelo majhne, diskretne spremembe v dednosti – mutacije. Mutacijska variabilnost kot dobavitelj materiala za selekcijo je naključna. Od tod tudi ime koncepta, ki ga je predlagal njegov kritik L.S. Berg (1922), "tihogeneza" evolucija, ki temelji na naključju.
Glavni ali celo edini gonilni dejavnik evolucije je naravna selekcija, ki temelji na selekciji (selekciji) naključnih in majhnih mutacij. Od tod tudi ime teorije - Selektogeneza.
Najmanjša razvijajoča se enota je populacija, ne posameznik, kot je domneval Charles Darwin. Zato je posebna pozornost namenjena proučevanju populacij kot strukturnih enot skupnosti: vrsta, čreda, jata.
Evolucija je postopna (gradacijska) in dolgoročna. Speciacija je pojmovana kot postopna zamenjava ene začasne populacije z zaporedjem naslednjih začasnih populacij.
Vrsta je sestavljena iz številnih podrejenih, hkrati morfološko, fiziološko in genetsko ločljivih, a ne reprodukcijsko izoliranih enot - podvrst, populacij (koncept široke politipske vrste).
Evolucija je po naravi divergentna (razhajanje značilnosti), t.j. en takson (sistematsko združevanje) lahko postane prednik več hčerinskih taksonov, vendar ima vsaka vrsta eno samo predniško vrsto, eno samo predniško populacijo.
Izmenjava alelov (pretok genov) je mogoča samo znotraj vrste. Zato je vrsta genetsko zaprt in celovit sistem.
Merila vrste ne veljajo za oblike, ki se razmnožujejo nespolno in partenogenetsko. To so lahko najrazličnejši prokarionti, nižji evkarionti brez spolnega procesa, pa tudi nekatere specializirane oblike višjih evkariontov, ki so sekundarno izgubili spolni proces (razmnožujejo se partenogenetsko).
Makroevolucija (tj. evolucija nad vrsto) poteka po poti mikroevolucije.
11. Evolucija je nepredvidljiva, tj. ima značaj, ki ni usmerjen h končnemu cilju.
V poznih 50. in zgodnjih 60. letih 20. stoletja so se pojavile dodatne informacije, ki kažejo na potrebo po reviziji nekaterih določb sintetične teorije. Vse večja je potreba po popravkih nekaterih njegovih določb.
Trenutno ostajajo veljavne 1., 2. in 3. teza teorije:
4. diplomska naloga se šteje kot neobvezna, ker evolucija se včasih lahko premika zelo hitro v skokih. Leta 1982 je bil v Dijonu (Francija) simpozij o stopnjah in oblikah speciacije. Pokazalo se je, da v V primeru poliploidije kromosomskih preureditev, ko se reproduktivna izolacija oblikuje skoraj takoj, se speciacija pojavi spazmodično. Kljub temu postopna speciacija s selekcijo majhnih mutacij v naravi nedvomno obstaja.
5. postulat je sporen, saj je znanih veliko vrst z omejenim arealom, znotraj katerega jih ni mogoče razdeliti na samostojne podvrste, reliktne vrste pa so lahko praviloma sestavljene iz ene populacije, usoda takih vrst pa je praviloma , kratkotrajno.
7. teza ostaja v veliki meri veljavna. Vendar pa so znani primeri uhajanja genov skozi ovire izolacijskih mehanizmov med posamezniki različnih vrst. Obstaja tako imenovani horizontalni prenos genov, na primer transdukcija - prenos bakterijskih genov iz ene vrste bakterij v drugo z okužbo z bakteriofagi. Obstajajo razprave o vprašanju horizontalnega prenosa genov. Število objav o tej problematiki narašča. Zadnji povzetek je predstavljen v monografiji R.B. Hasina "Spremenljivost genoma" (1984).
Transpozone, ki s selitvijo znotraj genoma povzročijo prerazporeditev zaporedja vključitve določenih genov, je treba obravnavati tudi z evolucijskega vidika.
8. diplomska naloga zahteva dopolnitev, saj ni jasno, kam uvrstiti organizme, ki se razmnožujejo nespolno in jih po tem kriteriju ne moremo uvrstiti med določene vrste.
9. teza je trenutno v reviziji, saj obstajajo dokazi, da lahko makroevolucija poteka tako skozi mikroevolucijo kot mimo tradicionalnih mikroevolucijskih poti.
10. teze - možnosti divergentnega izvora taksonov iz ene populacije prednikov (ali vrste) zdaj ne zanika nihče. Toda evolucija ni vedno različna. V naravi je pogosta oblika nastanka novih taksonov združevanje različnih, prej samostojnih, t.j. reproduktivno izolirane veje. Kombinacija različnih genomov in ustvarjanje novega uravnoteženega genoma poteka v ozadju delovanja naravne selekcije, ki zavrže nesposobne kombinacije genomov. V 30. letih je študent N.I. Vavilova V.I. Rybin je izvedel resintezo (obratno sintezo) gojene slive, katere izvor ni bil pojasnjen. V.A. Rybin je ustvaril svojo kopijo s hibridizacijo trnulj in češnje. Z resintezo so dokazali hibridogeni izvor nekaterih drugih vrst divjih rastlin. Botaniki menijo, da je hibridizacija eden od pomembnih načinov evolucije rastlin.
Revidirano je tudi 11. diplomsko delo. Ta problem je začel pritegniti posebno pozornost v zgodnjih dvajsetih letih našega stoletja, ko so dela N.I. Vavilov o homoloških serijah dedne variabilnosti. Opozoril je na obstoj določene smeri v variabilnosti organizmov in predlagal možnost njenega napovedovanja na podlagi analize serij homologne variabilnosti v sorodnih oblikah organizmov.
V dvajsetih letih so se pojavila dela domačega znanstvenika L.A. Berg, ki je izrazil misel, da je evolucija do neke mere vnaprej določena, kanalizirana v naravi, da obstajajo nekatere prepovedane poti evolucije, saj je število optimalnih rešitev v tem procesu očitno omejeno (teorija nomogeneze).
Na podlagi sodobnih konceptov lahko rečemo, da v evoluciji obstaja določena vektorizacija načinov preoblikovanja lastnosti in lahko do neke mere napovemo smer evolucije.
Sodobna teorija evolucije je torej nabrala ogromen arzenal novih dejstev in idej, vendar ni holistične teorije, ki bi lahko nadomestila sintetično teorijo evolucije, in to je stvar prihodnosti.
Po objavi glavnega dela Charlesa Darwina "Izvor vrst z naravno selekcijo" (1859) se je sodobna biologija močno oddaljila ne le od klasičnega darvinizma druge polovice 19. stoletja, temveč tudi od številnih določb sintetična teorija evolucije. Hkrati pa ni dvoma, da je glavna pot razvoja evolucijske biologije v skladu s tistimi smermi, ki jih je začrtal Darwin.
^ GENETSKI POLIMORFIZEM
Genetski polimorfizem razumemo kot stanje dolgotrajne raznolikosti genotipov, ko pogostnost tudi najbolj redkih genotipov v populacijah presega 1 %. Genetski polimorfizem se ohranja z mutacijami in rekombinacijami genskega materiala. Kot kažejo številne študije, je genetski polimorfizem zelo razširjen. Tako bo po teoretičnih izračunih potomstvo iz križanja dveh osebkov, ki se razlikujeta le v desetih lokusih, od katerih je vsak predstavljen s 4 možnimi aleli, vsebovalo približno 10 milijard osebkov z različnimi genotipi.
Večja kot je zaloga genetskega polimorfizma v določeni populaciji, lažje se prilagodi novemu okolju in hitreje poteka evolucija. Vendar pa je skoraj nemogoče oceniti število polimorfnih alelov s tradicionalnimi genetskimi metodami, saj se samo dejstvo prisotnosti gena v genotipu ugotovi s križanjem posameznikov z različnimi oblikami fenotipa, ki jih določa ta gen. Če vemo, kolikšen delež populacije predstavljajo posamezniki z različnimi fenotipi, lahko ugotovimo, koliko alelov je vključenih v nastanek določene lastnosti.
Od 60. let 20. stoletja se je metoda elektroforeze beljakovin (vključno z encimi) v gelu široko uporabljala za določanje genetskega polimorfizma. S to metodo je možno povzročiti gibanje proteinov v električnem polju, odvisno od njihove velikosti, konfiguracije in celotnega naboja, v različne dele gela, nato pa glede na lokacijo in število madežev, ki se pojavijo, preučevano snov je mogoče identificirati. Za oceno stopnje polimorfizma določenih proteinov v populacijah se običajno preučuje približno 20 ali več lokusov, nato pa se matematično določi število alelnih genov ter razmerje med homo- in heterozigoti. Raziskave kažejo, da so nekateri geni ponavadi monomorfni, medtem ko so drugi izjemno polimorfni.
Obstajata prehodni in uravnoteženi polimorfizem, ki je odvisen od selektivne vrednosti genov in pritiska naravne selekcije.
^ Prehodni polimorfizem se pojavi v populaciji, ko je alel, ki je bil nekoč pogost, zamenjan z drugimi aleli, ki dajejo njihovim nosilcem večjo sposobnost (multipli alelizem). Pri prehodnem polimorfizmu opazimo smerni premik v odstotku oblik genotipa. Prehodni polimorfizem je glavna pot evolucije, njena dinamika. Primer prehodnega polimorfizma je lahko pojav industrijskega mehanizma. Tako so se zaradi onesnaženosti zraka v industrijskih mestih Anglije v zadnjih sto letih pojavile temne oblike pri več kot 80 vrstah metuljev. Na primer, če so imeli pred letom 1848 brezovi molji bledo kremno barvo s črnimi pikami in posameznimi temnimi pikami, so se leta 1848 v Manchestru pojavile prve temno obarvane oblike, do leta 1895 pa je 98% moljev postalo temno obarvanih. Do tega je prišlo zaradi sajanja drevesnih debel in selektivnega objedanja svetlobnih vešč s strani drozgov in robinov. Kasneje je bilo ugotovljeno, da temno obarvanost telesa moljev povzroča mutirani melanistični alel.
^ Uravnotežen polimorfizem x značilna je odsotnost premika v številčnih razmerjih različnih oblik in genotipov v populacijah, ki se nahajajo v stabilnih okoljskih razmerah. V tem primeru odstotek oblik ostaja enak iz generacije v generacijo ali pa niha okoli neke stalne vrednosti. V nasprotju s prehodnim je uravnoteženi polimorfizem statična evolucija. I.I. Schmalhausen (1940) ga je imenoval ravnotežni heteromorfizem.
Primer uravnoteženega polimorfizma je prisotnost dveh spolov pri monogamnih živalih, saj imajo enake selektivne prednosti. Njihovo razmerje v populacijah je 1:1. Pri poligamiji se lahko selektivna vrednost predstavnikov različnih spolov razlikuje, nato pa so predstavniki enega spola uničeni ali v večji meri kot posamezniki drugega spola izključeni iz razmnoževanja. Drug primer so človeške krvne skupine po sistemu ABO. Pri tem se pogostost različnih genotipov v različnih populacijah lahko razlikuje, vendar v vsaki posamezni populaciji ostaja nespremenjena iz generacije v generacijo. To je razloženo z dejstvom, da noben genotip nima selektivne prednosti pred drugimi. Torej, čeprav imajo moški s prvo krvno skupino, kot kažejo statistični podatki, daljšo pričakovano življenjsko dobo kot moški z drugimi krvnimi skupinami, je pri njih večja verjetnost, da razvijejo razjedo na dvanajstniku, ki lahko v primeru perforacije povzroči smrt.
Genetsko ravnovesje v populacijah je lahko porušeno zaradi pritiska spontanih mutacij, ki se pojavljajo z določeno pogostostjo v vsaki generaciji. Vztrajnost ali odprava teh mutacij je odvisna od tega, ali jim daje prednost naravna selekcija ali se upira. Če sledimo usodi mutacij v določeni populaciji, lahko govorimo o njeni prilagoditveni vrednosti. Slednja je enaka 1, če je selekcija ne izključuje in ne preprečuje njenega širjenja. V večini primerov je adaptivna vrednost mutantnih genov manjša od 1, in če se mutanti popolnoma ne morejo razmnoževati, potem je nič. Tovrstne mutacije naravna selekcija zavrne. Vendar pa lahko isti gen večkrat mutira, kar kompenzira njegovo eliminacijo, ki jo povzroči selekcija. V takšnih primerih je mogoče doseči ravnovesje, kjer sta pojav in izginotje mutiranih genov uravnotežena. Primer je srpastocelična anemija, ko dominantni mutirani gen v homozigotu povzroči zgodnjo smrt telesa, vendar so heterozigoti za ta gen odporni na malarijo. Na območjih, kjer je malarija pogosta, obstaja uravnotežen polimorfizem v genu za anemijo srpastih celic, saj skupaj z izločanjem homozigotov obstaja nasprotna selekcija v korist heterozigotov. Kot rezultat večvektorske selekcije se genotipi ohranjajo v genskem skladu populacij v vsaki generaciji, kar zagotavlja ustreznost organizmov ob upoštevanju lokalnih razmer. Poleg gena za srpaste celice obstajajo v človeški populaciji številni drugi polimorfni geni, za katere se domneva, da povzročajo pojav heteroze
Recesivne mutacije (vključno s škodljivimi), ki se fenotipsko ne manifestirajo pri heterozigotih, se lahko kopičijo v populacijah do višje ravni kot škodljive dominantne mutacije.
Genetski polimorfizem je predpogoj za neprekinjen razvoj. Zahvaljujoč temu lahko v spreminjajočem se okolju vedno obstajajo genetske različice, vnaprej prilagojene tem pogojem. V populaciji diploidnih dvodomnih organizmov je lahko velika rezerva genetske variabilnosti shranjena v heterozigotnem stanju, ne da bi se fenotipsko manifestirala. Raven slednjega je očitno lahko še višja pri poliploidnih organizmih, pri katerih se lahko za fenotipsko manifestiranim normalnim alelom skriva ne en, ampak več mutantnih alelov.
^ GENSKA OBREMENITEV
Genetska fleksibilnost (ali plastičnost) populacij se doseže s procesom mutacije in kombinacijske variabilnosti. In čeprav je evolucija odvisna od nenehne prisotnosti genetske variacije, je ena od njenih posledic pojav slabo prilagojenih osebkov v populacijah, zaradi česar je pripravljenost populacij vedno nižja od tiste, ki je značilna za optimalno prilagojene organizme. To zmanjšanje povprečne telesne pripravljenosti populacije zaradi posameznikov, katerih telesna pripravljenost je pod optimalno, se imenuje genetska obremenitev. Kot je zapisal slavni angleški genetik J. Haldane, ki je opisal genetsko obremenitev: "To je cena, ki jo je populacija prisiljena plačati za pravico do razvoja." Bil je prvi, ki je raziskovalce opozoril na obstoj genetske obremenitve, sam izraz »genetska obremenitev« pa je v 40. letih 20. stoletja uvedel G. Miller.
Genetska obremenitev v širšem smislu je kakršno koli zmanjšanje (dejansko ali potencialno) sposobnosti populacije zaradi genetske variabilnosti. Kvantificiranje genetske obremenitve in določanje njenega resničnega vpliva na sposobnost populacije je težka naloga. Po predlogu F. G. Dobzhansky (1965) se posamezniki, katerih sposobnost je več kot dve standardni deviaciji (-2a) pod povprečno sposobnostjo heterozigotov, štejejo za nosilce genetskega tovora.
Običajno ločimo tri vrste genetske obremenitve: mutacijsko, substitucijsko (prehodno) in uravnoteženo. Celotno genetsko obremenitev sestavljajo te tri vrste obremenitev. Mutacijska obremenitev - to je delež celotne genetske obremenitve, ki nastane zaradi mutacij. Ker pa je večina mutacij škodljivih, je naravna selekcija usmerjena proti takim alelom in njihova pogostnost je nizka. V populacijah se ohranjajo predvsem zaradi na novo nastalih mutacij in heterozigotnih prenašalcev.
Genetska obremenitev, ki nastane pri dinamičnih spremembah genskih frekvenc v populaciji v procesu zamenjave enega alela z drugim, se imenuje nadomestni (ali prehodni) tovor. Takšna zamenjava alelov se običajno pojavi kot odgovor na neko spremembo okoljskih razmer, ko prej neugodni aleli postanejo ugodni in obratno (primer bi bil pojav industrijskega mehanizma metuljev na ekološko neugodnih območjih). V tem primeru se frekvenca enega alela zmanjšuje, ko se frekvenca drugega povečuje.
^ Uravnotežen (stabilen) polimorfizem se zgodi, ko se številne lastnosti vzdržujejo na relativno konstantnih ravneh z uravnoteženo selekcijo. Hkrati se zaradi uravnotežene selekcije, ki deluje v nasprotnih smereh, v populacijah ohranita dve ali več alej katerega koli lokusa in posledično se ohranijo različni genotipi in fenotipi. Primer je srpasta celica. Pri tem je selekcija usmerjena proti mutantnemu alelu, ki je v homozigotnem stanju, hkrati pa deluje v korist heterozigota in ga ohranja. Stanje uravnotežene obremenitve je mogoče doseči v naslednjih situacijah: 1) selekcija daje prednost danemu alelu na eni stopnji ontogeneze in je usmerjena proti njemu na drugi; 2) selekcija daje prednost ohranitvi alela pri posameznikih enega spola in deluje proti njemu pri posameznikih drugega spola; 3) znotraj istega alela različni genotipi omogočajo organizmom uporabo različnih ekoloških niš, kar zmanjšuje tekmovalnost in posledično slabi eliminacijo; 4) v subpopulacijah, ki zasedajo različne habitate, je selekcija naklonjena različnim alelom; 5) selekcija daje prednost ohranitvi alela, medtem ko je redek in je usmerjen proti njemu, ko se pojavlja pogosto.
Veliko poskusov je bilo oceniti dejansko gensko obremenitev v človeški populaciji, vendar se je to izkazalo za zelo težko nalogo. O njem je mogoče posredno soditi po stopnji prenatalne umrljivosti in rojstva otrok z nekaterimi oblikami razvojnih nepravilnosti, zlasti od staršev v sorodstvenih zakonih, še bolj pa v incestu.
Literatura:
1. Abrikosov G.G., Bekker Z.G. in drugi Tečaj zoologije v dveh zvezkih. Zvezek I.- Zoologija nevretenčarjev. 7. izdaja. Založnik: "Višja šola", M., 1966.-552 str.
2. Zaponka, John. Živalski hormoni (iz angleščine prevedla M.S. Morozova). Založnik: Mir, 1986.-85(1) str.
3. Beklemišev V.N. Osnove primerjalne anatomije nevretenčarjev. Založnik: Sov. Znanost, M., 1944.-489p.
4. Volkova O.V., Pekarsky M.I. Embriogeneza in s starostjo povezana histologija človeških notranjih organov. Založnik: "Medicina", M, 1976. 45s.
5. Gurtovoy N.N., Matveev B.S., Dzerzhinsky F.Ya. Praktična zootomija vretenčarjev. Dvoživke in plazilci./Ed. B.S. Matveeva in N.N. Gurtovoy. Založnik: "Višja šola", M., 1978. - 406 str.
6. Gaivoronsky I.V. Normalna človeška anatomija: učbenik. v 2 zvezkih / I.V. Gaivoroksky - 3. izd., popravljeno. – St. Petersburg: Spetslit, 2003, letnik 1 – 2003. – 560 str., letnik 2 – 2003. – 424 str.
7.Histologija (uvod v patologijo). Učbenik za študente višje medicine. izobraževalne ustanove./Ur. E.G.Ulumbekova, Yu.A. Chelysheva. Založnik: "GEOTAR", M., 1997. - 947 str.
8. Zussman, M. Razvojna biologija./ Ed. S.G. Vasetsky. per. iz angleščine Založnik: "Svet". M. 1977-301s.
9. Levina S.E. Eseji o razvoju spola v zgodnji ontogenezi višjih vretenčarjev. Založnik: "Science". M., 1974.-239p.
10. Leibson L.G. Glavne značilnosti strukturnega in funkcionalnega razvoja endokrinega sistema vretenčarjev. Dnevnik evol. Biokemija in fiziologija, 1967, letnik 3., št. 6, str. 532 – 544.
11. Lukin E.I. Zoologija: učbenik za študente zooinženirskih in zoo-veterinarskih univerz in fakultet. – 2. izd., predelana. in dodatno – Založnik: “Višja šola”, 1981, M. - 340 str.
12. Naumov S.P. Zoologija vretenčarjev. Založnik: "Razsvetljenje", M., 1982.-464 str.
13. Talyzin F.F., Ulisova T.N. Gradivo za primerjalno anatomijo organskih sistemov vretenčarjev. Učbenik za študente. M., 1974.-71 str.
14. Fiziologija človeka in živali (splošna in evolucijsko-ekološka), v 2 delih. Ed. Kogan A.B. Založnik: "Višja šola". M. 1984, I. del - 360 str., II. del - 288 str.
15. Šmalgauzen I.I. Osnove primerjalne anatomije. Država založba biol. in medicinsko literaturo. M., 1935.-924 str.
O nekaterih vprašanjih evolucije
2. izdaja, razširjena
Sestavil: Khojayan A.B., Krasnova L.A., Fedorenko N.N.
Dostavljeno za zaposlitev 07/05/09. Podpisano v tisk 05.07.09.-Format 60x99.
Tipičen papir Št. 1. Ofsetni tisk. Slušalke so zamaknjene. Pogojno pečica l. 2.0.
Akademska ur. l. 1.2. Naročilo 2087. Naklada 100.
Državna medicinska akademija Stavropol.
355017, Stavropol, ul. Mira, 310.
Nastajanje možganov pri zarodkih vseh vretenčarjev se začne s pojavom oteklin - možganskih veziklov - na sprednjem koncu nevralne cevi. Najprej so trije, nato pa pet. Iz prednjih možganov se nato oblikujejo prednji možgani in diencefalon, iz srednjega - mezencefalon, iz zadnjega pa - mali možgani in podolgovata medula. Slednji brez ostre meje prehaja v hrbtenjačo
V nevralni cevi je votlina - neurocoel, ki med nastankom petih možganskih veziklov tvori podaljške - možganske prekate (v teh predelih možganov so 4) dno (baza) in a ločimo streho (plašč). Streha se nahaja zgoraj - in dno se nahaja pod ventrikli.
Možganska snov je heterogena – predstavljata jo siva in bela snov. Siva je skupek nevronov, bela pa nastane iz odrastkov nevronov, prekritih z maščobo podobno snovjo (mielinska ovojnica), ki daje možganski snovi belo barvo. Plast sive snovi na strehi katerega koli dela možganov se imenuje skorja.
Čutilni organi igrajo pomembno vlogo pri razvoju živčnega sistema. Koncentracija čutnih organov na sprednjem koncu telesa je bila tista, ki je določila progresivni razvoj glave nevralne cevi. Menijo, da je sprednji možganski mehurček nastal pod vplivom vohalnih, srednjih - vizualnih in posteriornih - slušnih receptorjev.
ribe
Prednji možgani majhna, ni razdeljena na poloble, ima samo en ventrikel. Njena streha ne vsebuje živčnih elementov, ampak jo tvori epitelij. Nevroni so koncentrirani na dnu ventrikla v striatumu in v vohalnih režnjih, ki se raztezajo pred sprednjimi možgani. V bistvu prednji možgani delujejo kot vohalni center.
Srednji možgani je najvišje regulativno in integrativno središče. Sestavljen je iz dveh optičnih režnjev in je največji del možganov. Ta vrsta možganov, kjer je najvišji regulacijski center srednji možgan, se imenuje ihtiopsidpim.
diencefalon sestavljena iz strehe (talamus) in dna (hipotalamus) je povezana s hipotalamusom, pinealna žleza pa je povezana s talamusom.
Mali možgani pri ribah je dobro razvit, saj so njihova gibanja zelo raznolika.
Medulla oblongata brez ostre meje prehaja v hrbtenjačo in v njej so skoncentrirani prehranjevalni, vazomotorični in dihalni centri.
»1. stopnja. Tvorba centralnega živčnega sistema v obliki cevi se najprej pojavi pri živalih hordata. Pri nižjih strunastih, na primer suličniku, je nevralna cev ohranjena vse življenje, pri višjih strunastih - vretenčarjih - je v embrionalni fazi na hrbtni strani zarodka položena nevralna plošča, ki se pogrezne pod kožo in se zloži v plast; cev.
Stopnja 2. Pri vretenčarjih je nevralna cev razdeljena na možgane in hrbtenjačo. Nevralna cev v embrionalni fazi razvoja tvori v sprednjem delu tri otekline - tri možganske mehurčke, iz katerih se razvijejo deli možganov: iz sprednjega mešička nastaneta prednji del možganov in diencefalon, srednji mešiček preide v srednje možgane, posteriorni vezikel tvorita cerebelum in medulla oblongata. Teh pet predelov možganov je značilnih za vse vretenčarje.
Faza 3. Za nižje vretenčarje - ribe in dvoživke - je značilna prevlada srednjih možganov nad drugimi deli. Samo pri hrustančnih ribah morskega psa so zaradi hitrega gibanja razviti mali možgani, močno razvit voh pa je povzročil povečanje prednjih možganov, ki postanejo središče za obdelavo vohalnih signalov.
Faza 4. Pri dvoživkah se prednji del možganov rahlo poveča in na strehi hemisfer se oblikuje tanka plast živčnih celic - primarni medularni obok (archipallium), starodavna skorja. Poleg arhipalija so pri dvoživkah okrepljene povezave med prednjimi in srednjimi možgani.
Stopnja 5. Pri plazilcih se prednji del možganov znatno poveča zaradi kopičenja živčnih celic - progastih teles - na dnu prednjih možganov. Večino strehe hemisfer zavzema starodavna skorja. Prvič pri plazilcih se pojavi zametek nove skorje - neopalija. Hemisfere prednjih možganov se plazijo na druge dele, zaradi česar nastane ovinek v predelu diencefalona. Od starodavnih plazilcev so možganske hemisfere postale največji del možganov.
Struktura možganov ptic in plazilcev ima veliko skupnega. Na strehi možganov je primarna skorja, srednji možgani so dobro razviti. Vendar se pri pticah v primerjavi s plazilci skupna masa možganov in relativna velikost prednjih možganov povečata. Veliki optični režnji srednjih možganov kažejo na povečano vlogo vida pri vedenju ptic. Mali možgani so veliki in imajo nagubano strukturo.
Pomemben del obeh polobel prednjih možganov pri pticah, tako kot pri plazilcih, tvori striatum - izrastke dna prednjih možganov.
Stopnja 6. Pri sesalcih prednji možgani dosežejo največjo velikost in kompleksnost. Večino možganske snovi sestavlja nova skorja - sekundarni medularni obok ali neopalij. Sestavljen je iz živčnih celic in vlaken, ki so razporejena v več plasteh. Neokorteks možganskih hemisfer služi kot središče višjega živčnega delovanja.
Vmesni in srednji del možganov pri sesalcih sta majhna. Širječe hemisfere prednjih možganov jih pokrivajo in stiskajo pod seboj. Pri primatih hemisfere sprednjih možganov pokrivajo tudi male možgane, pri človeku pa podolgovato medulo. Nekateri sesalci imajo gladke možgane brez utorov ali vijug, večina sesalcev pa ima utore in vijuge v možganski skorji, ki nastanejo, ko skorja raste. Največje žlebičaste tvorbe so pri kitih, najmanjše pri žužkojedih in netopirjih.
Faza 7. Pojav žlebov in konvolucij se pojavi zaradi rasti možganov z omejenimi dimenzijami lobanje. Zdi se, da so možgani vtisnjeni v kostne stene lobanje, možganske membrane pa so stisnjene. Nadaljnja rast skorje vodi do pojava zlaganja v obliki utorov in zvitkov. V možganski skorji vseh sesalcev so jedrske cone analizatorjev, tj. polja primarne kortikalne analize."
Deryagina M.A., Evolucijska antropologija: biološki in kulturni vidiki, M., Založba "URAO", 2003, str. 148-150.
Tvorba možganov pri vseh vretenčarjih se začne s tvorbo treh oteklin ali možganskih veziklov na sprednjem koncu nevralne cevi: sprednjega, srednjega in zadnjega. Nato je sprednji medularni vezikel razdeljen s prečno zožitvijo na dva dela. Prvi od njih tvori sprednji del možganov, ki pri večini vretenčarjev tvori možganske hemisfere. Na zadnji strani se razvije sprednji medularni mehur diencefalon. Srednji možganski vezikel se ne deli in se popolnoma preoblikuje v srednji možgani. Posteriorni medularni vezikel je prav tako razdeljen na dva dela: v sprednjem delu tvori zadnji možgani ali mali možgani, in iz zadnjega dela nastane medulla oblongata, ki brez ostre meje prehaja v hrbtenjača.
V procesu nastajanja petih možganskih veziklov tvori votlina nevralne cevi niz podaljškov, ki se imenujejo možganski ventrikli. Votlina prednjih možganov se imenuje stranski prekat, vmesni - tretji prekat, medulla oblongata - četrti prekat, srednji možgani - Sylvian kanal, ki povezuje 3. in 4. prekat. Zadnji možgani nimajo votline.
V vsakem delu možganov so streha, ali haljo in dno, ali bazo. Streho sestavljajo deli možganov, ki ležijo nad prekati, dno pa deli pod komorami.
Snov možganov je heterogena. Temna področja so siva snov, svetla pa bela snov. Bela snov je skupek živčnih celic z mielinsko ovojnico (številni lipidi, ki dajejo belkasto barvo). Siva snov je skupek živčnih celic med nevroglialnimi elementi. Plast sive snovi na površini strehe katerega koli dela možganov se imenuje skorja.
Pri vseh vretenčarjih so možgani sestavljeni iz petih delov, ki so razporejeni v istem zaporedju. Vendar stopnja njihovega razvoja pri predstavnikih različnih razredov ni enaka. Te razlike so posledica filogenije.
Obstajajo tri vrste možganov: ihtiopsid, sauropsid in sesalec.
TO ihtipsidnega tipa možgani vključujejo možgane rib in dvoživk. Je vodilni del možganov, središče refleksne aktivnosti.
Ribji možgani ima primitivno strukturo, ki se izraža v majhni velikosti možganov kot celote in slabem razvoju sprednjega dela. Sprednji možgani so majhni in niso razdeljeni na poloble. Streha prednjih možganov je tanka. Pri koščenih ribah ne vsebuje živčnega tkiva. Njen glavni del tvori dno, kjer živčne celice tvorijo dve skupini – striatum. Dva vohalna režnja segata naprej od prednjih možganov. Prednji možgani ribe opravlja funkcijo centra za voh.
Diencephalon rib od zgoraj pokrit s sprednjim in srednjim delom. Iz njegove strehe se razteza izrastek - pinealna žleza - lijak s sosednjo hipofizo in vidnimi živci.
Srednji možgani- najbolj razvit del ribjih možganov. To je vidno središče rib in je sestavljeno iz dveh optičnih reženj. Na površini strehe je plast sive snovi (lubje). To je najvišji del ribjih možganov, saj prihajajo signali vseh dražljajev in tu nastajajo odzivni impulzi. Mali možgani rib so dobro razviti, saj so gibi rib raznoliki.
Medulla oblongata pri ribah ima visoko razvite visceralne režnje in je povezan z močnim razvojem okusnih organov.
Možgani dvoživk ima številne progresivne spremembe, povezane s prehodom na življenje na kopnem, ki se izražajo v povečanju skupnega volumna možganov in razvoju njegovega sprednjega dela. Hkrati je prednji del možganov razdeljen na dve polobli. Streha prednjih možganov je sestavljena iz živčnega tkiva. Na dnu prednjih možganov leži striatum. Vohalni režnji so ostro omejeni od hemisfer. Prednji del možganov ima še vedno pomen le centra za voh.
diencefalon jasno viden od zgoraj. Njegovo streho tvori privesek - epifiza, in dno - hipofiza.
Srednji možgani manjša od rib. Hemisfere srednjih možganov so dobro definirane in prekrite s skorjo. To je vodilni oddelek centralnega živčnega sistema, ker Tu se prejete informacije analizirajo in generirajo odzivni impulzi. Ohranja pomen vizualnega središča.
Mali možgani je slabo razvit in ima videz majhnega prečnega grebena na sprednjem robu romboidne fose medule oblongate. Slab razvoj malih možganov ustreza enostavnim gibanjem dvoživk.
TO sauropsidnega tipa možganov vključujejo možgane plazilcev in ptic.
Pri plazilcih opazimo nadaljnje povečanje volumna možganov. Prednji del možganov postane največji del. Preneha biti le vohalni center in postane vodilni oddelek centralnega živčnega sistema zaradi dna, kjer se razvije striatum. Prvič v procesu evolucije se na površini možganov pojavijo živčne celice oziroma skorja, ki ima primitivno strukturo (tri plasti) in se imenuje starodavna skorja - arheokorteks.
diencefalon je zanimiv za strukturo hrbtnega dodatka - parietalnega organa ali parietalnega očesa, ki doseže svoj najvišji razvoj pri kuščarjih, pridobi strukturo in funkcijo organa vida.
Srednji možgani se zmanjša, izgubi pomen vodilnega oddelka in zmanjša se njegova vloga vizualnega središča.
Mali možgani razmeroma bolje razviti kot pri dvoživkah.
Za ptičje možgane za katerega je značilno nadaljnje povečanje celotnega volumna in ogromna velikost prednjih možganov, ki pokriva vse druge dele razen malih možganov. Povečanje prednjih možganov, ki so tako kot pri plazilcih vodilni del možganov, se zgodi na račun dna, kjer se močno razvije striatum. Streha prednjih možganov je slabo razvita in ima majhno debelino. Korteks se ne razvija naprej in se celo podvrže obratnemu razvoju - stranski del skorje izgine.
diencefalon majhna , Pinealna žleza je slabo razvita, hipofiza je dobro izražena.
IN srednji možgani vidni režnji so razviti, saj vid igra vodilno vlogo v življenju ptic.
Mali možgani doseže ogromne velikosti in ima zapleteno strukturo. Ima srednji del in stranske izbokline. Razvoj malih možganov je povezan z letom.
Proti tipu mlečnih možganov vključujejo možgane sesalcev.
Evolucija možganov je šla v smeri razvoja strehe prednjih možganov in hemisfer, s čimer se je povečala površina prednjih možganov zaradi vijug in utorov skorje.
Na celotni površini strehe se pojavi plast sive snovi – pravi lubje. To je popolnoma nova struktura, ki nastane med evolucijo živčnega sistema. Pri nižjih sesalcih je površina skorje gladka, pri višjih sesalcih pa tvori številne vijuge, ki močno povečajo njeno površino. Prednji možgani postane pomemben vodilni oddelek možganov zaradi razvoja skorje, ki je značilna za mamarni tip. Zelo razviti so tudi vohalni režnji, saj so pri mnogih sesalcih čutni organ.
diencefalon ima značilne dodatke - pinealno žlezo, hipofizo. Srednji možgani zmanjšana velikost. Njena streha ima poleg vzdolžne brazde tudi prečno. Zato se namesto dveh hemisfer (optičnih režnjev) oblikujejo štirje tuberkuli. Sprednji tuberkuli so povezani z vidnimi receptorji, zadnji pa s slušnimi.
Mali možgani se razvija postopoma, kar se izraža v močnem povečanju velikosti organa in njegove kompleksne zunanje in notranje strukture.
V podolgovati meduli je ob straneh pot živčnih vlaken, ki vodijo do malih možganov, na spodnji površini pa so vzdolžni grebeni (piramide).
12. Vrste, oblike in pravila evolucije skupin (na kratko pravila makroevolucije)
Makroevolucija je niz evolucijskih transformacij, ki se dogajajo na ravni nadspecifičnih taksonov. Nadspecifični taksoni (rodovi, družine, redovi, razredi) so zaprti genetski sistemi. [Za označevanje mehanizmov nastanka višjih taksonov (oddelkov, tipov) je J. Simpson uvedel izraz »megaevolucija«.] Prenos genov iz enega zaprtega sistema v drugega je nemogoč ali malo verjeten. Tako prilagoditvene lastnosti, ki je nastala v enem zaprtem taksonu, ni mogoče prenesti na drug zaprti takson. Zato med makroevolucijo nastanejo pomembne razlike med skupinami organizmov. Zato lahko na makroevolucijo gledamo kot na razvoj zaprtih genetskih sistemov, ki v naravnih razmerah ne morejo izmenjevati genov.
1. Pravilo nepovratnosti evolucije, ali Dollotov princip (Louis Dollot, belgijski paleontolog, 1893): izginila lastnost se ne more znova pojaviti v prejšnji obliki. Na primer, sekundarni vodni mehkužci in vodni sesalci niso obnovili dihanja škrg.
2. Pravilo porekla od nespecializiranih prednikov, ali Copov princip (Edward Cope, ameriški paleontolog-zoolog, 1904): nova skupina organizmov nastane iz nespecializiranih prednikov. Na primer, nespecializirani žužkojedi (kot so sodobni tenreki) so povzročili vse sodobne placentne sesalce.
3. Pravilo progresivne specializacije, ali Deperejev princip (C. Depere, paleontolog, 1876): skupina, ki je stopila na pot specializacije, bo v svojem nadaljnjem razvoju sledila poti vse globlje specializacije. Sodobni specializirani sesalci (Chiroptera, plavutonožci, kiti) se bodo najverjetneje razvijali po poti nadaljnje specializacije.
4. Adaptivno radiacijsko pravilo, ali načelo Kovalevsky-Osborne (V.O. Kovalevsky, Henry Osborne, ameriški paleontolog): skupina, v kateri se pojavi brezpogojno napredujoča lastnost ali skupek takšnih lastnosti, povzroči številne nove skupine, ki tvorijo številne nove ekološke niše in zahajajo celo v druge habitate. Iz primitivnih placentnih sesalcev so na primer nastale vse sodobne evolucijsko-ekološke skupine sesalcev.
5. Pravilo integracije bioloških sistemov, ali Schmalhausenov princip (I.I. Shmalhausen): nove, evolucijsko mlade skupine organizmov absorbirajo vse evolucijske dosežke skupin prednikov. Na primer, sesalci so uporabili vse evolucijske dosežke prednikov: mišično-skeletni sistem, čeljusti, parne okončine, glavne dele centralnega živčnega sistema, embrionalne membrane, popolne izločevalne organe (medenične ledvice), različne derivate povrhnjice itd.
6. Pravilo menjave faze, ali načelo Severtsov-Schmalhausen (A.N. Severtsov, I.I. Shmalhausen): različni mehanizmi evolucije se naravno zamenjujejo. Na primer, alomorfoze prej ali slej postanejo aromorfoze in na podlagi aromorfoz nastanejo nove alomorfoze.
Poleg pravila za menjavanje faz je J. Simpson uvedel pravilo za menjavanje hitrosti evolucije; Na podlagi hitrosti evolucijskih preobrazb je ločil tri tipe evolucije: braditelično (počasen tempo), horotelično (srednje tempo) in itachitelično (hiter tempo).
II. VPRAŠANJA ZA KOLOKVIJ IN IZPITE IZ GENETIKE.
1. Genetika(iz grščine γενητως - ki izvira od nekoga) - znanost o zakonih dednosti in variabilnosti. Glede na predmet študija je razvrščena genetika rastlin, živali, mikroorganizmov, ljudi in drugih; odvisno od metod, ki se uporabljajo v drugih disciplinah – molekularni genetiki, okoljski genetiki in drugih. Dednost običajno opredeljena kot sposobnost organizmov za razmnoževanje lastne vrste, kot lastnost starševskih osebkov, da svoje značilnosti in lastnosti prenašajo na svoje potomce. Ta izraz opredeljuje tudi podobnost sorodnih posameznikov med seboj.
Genetske raziskovalne metode:
Hibridološka metoda je prvi razvil in uporabil G. Mendel v letih 1856-1863. za preučevanje dedovanja lastnosti in je od takrat glavna metoda genetskih raziskav. Vključuje sistem križanj vnaprej izbranih staršev, ki se razlikujejo po enem, dveh ali treh alternativnih znakih, katerih dedovanje se proučuje. Glede na stopnjo in naravo manifestacije proučevanih lastnosti se izvede temeljita analiza hibridov prve, druge, tretje in včasih naslednjih generacij. Ta metoda je pomembna pri žlahtnjenju rastlin in živali. Vključuje tudi t.i metoda rekombinacije , ki temelji na pojavu prečkanje- izmenjava identičnih regij v kromatidah homolognih kromosomov v profazi I mejoze. Ta metoda se pogosto uporablja za sestavljanje genetskih zemljevidov, pa tudi za ustvarjanje rekombinantnih molekul DNK, ki vsebujejo genetske sisteme različnih organizmov.
Monosomska metoda omogoča ugotavljanje, v katerem kromosomu so lokalizirani ustrezni geni, in v kombinaciji z metodo rekombinacije določanje lokacije genov na kromosomu.
Genealoška metoda - ena od hibridoloških možnosti. Uporablja se pri preučevanju dedovanja lastnosti z analizo rodovnikov, ob upoštevanju njihove manifestacije pri živalih sorodnih skupin v več generacijah. Ta metoda se uporablja pri preučevanju dednosti pri ljudeh in živalih, katerih neplodnost je določena glede na vrsto.
Dvojna metoda uporablja se pri preučevanju vpliva določenih okoljskih dejavnikov in njihove interakcije z genotipom posameznika, pa tudi za ugotavljanje relativne vloge genotipske in modifikacijske variabilnosti v splošni variabilnosti lastnosti. Dvojčki so potomci, rojeni v istem leglu samskih domačih živali (govedo, konji itd.).
Obstajata dve vrsti dvojčkov - enojajčni (identični), ki imajo enak genotip, in neidentični (brajski), ki izhajajo iz ločeno oplojenih dveh ali več jajčec.
Metoda mutacije (mutageneza) omogoča ugotavljanje narave vpliva mutagenih dejavnikov na genetski aparat celice, DNA, kromosome in na spremembe značilnosti ali lastnosti. Mutageneza se uporablja pri žlahtnjenju kmetijskih rastlin in v mikrobiologiji za ustvarjanje novih sevov bakterij. Našel je uporabo pri izbiri sviloprejk.
Metoda populacijske statistike uporablja pri proučevanju pojavov dednosti v populacijah. Ta metoda omogoča ugotavljanje pogostosti dominantnih in recesivnih alelov, ki določajo določeno lastnost, pogostost dominantnih in recesivnih homozigotov in heterozigotov, dinamiko genetske strukture populacije pod vplivom mutacij, izolacije in selekcije. Metoda je teoretična osnova sodobne selekcije živali.
Fenogenetska metoda nam omogoča, da ugotovimo stopnjo vpliva genov in okoljskih pogojev na razvoj proučevanih lastnosti in značilnosti v ontogenezi. Spremembe v hranjenju in vzdrževanju živali vplivajo na naravo manifestacije dedno določenih lastnosti in lastnosti.
Sestavni del vsake metode je statistična analiza – biometrična metoda . Predstavlja niz matematičnih tehnik, ki omogočajo ugotavljanje stopnje zanesljivosti dobljenih podatkov in ugotavljanje verjetnosti razlik med kazalniki poskusne in kontrolne skupine živali. Sestavni del biometrije sta zakon regresije in statistični zakon dednosti, ki ju je postavil F. Galton.
Pogosto se uporablja v genetiki metoda modeliranja z uporabo računalnika preučevati dedovanje kvantitativnih lastnosti v populacijah, vrednotiti selekcijske metode - množična selekcija, selekcija živali po selekcijskih indeksih. Ta metoda je našla posebno široko uporabo na področju genskega inženiringa in molekularne genetike.




